

Fertilization Mechanisms in Flowering Plants
- PMID: 26859271
- PMCID: PMC4934421
- DOI: 10.1016/j.cub.2015.12.032
Fertilization Mechanisms in Flowering Plants
Abstract
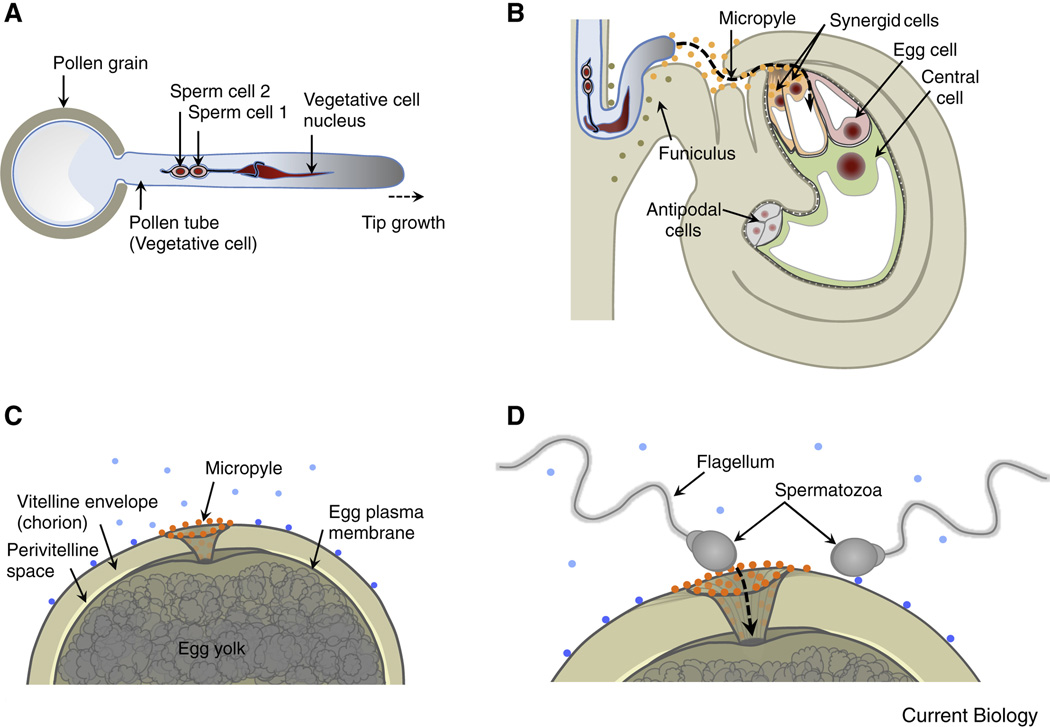
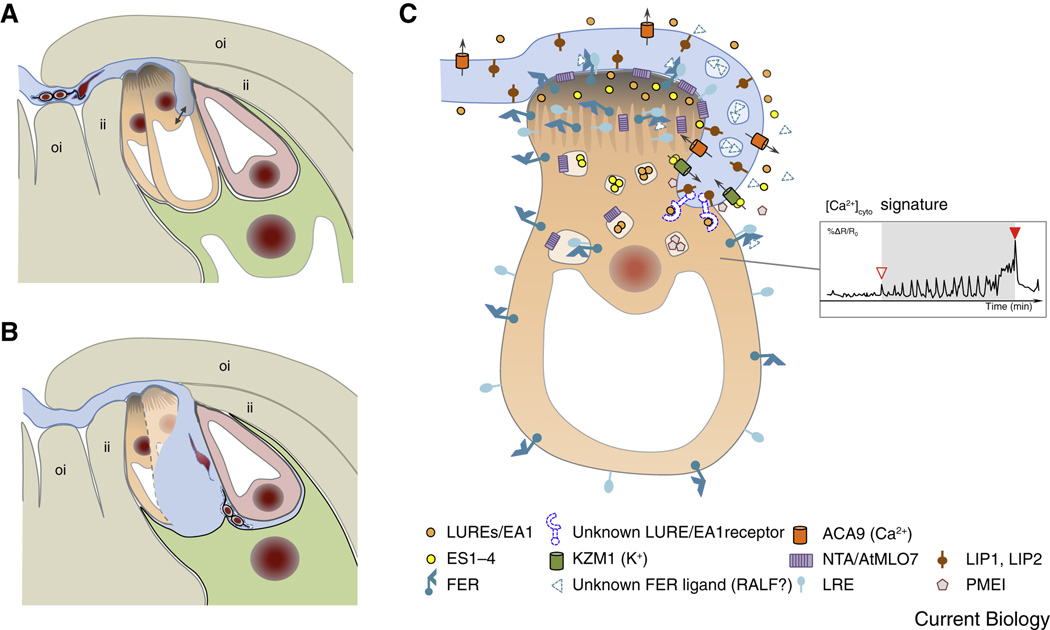
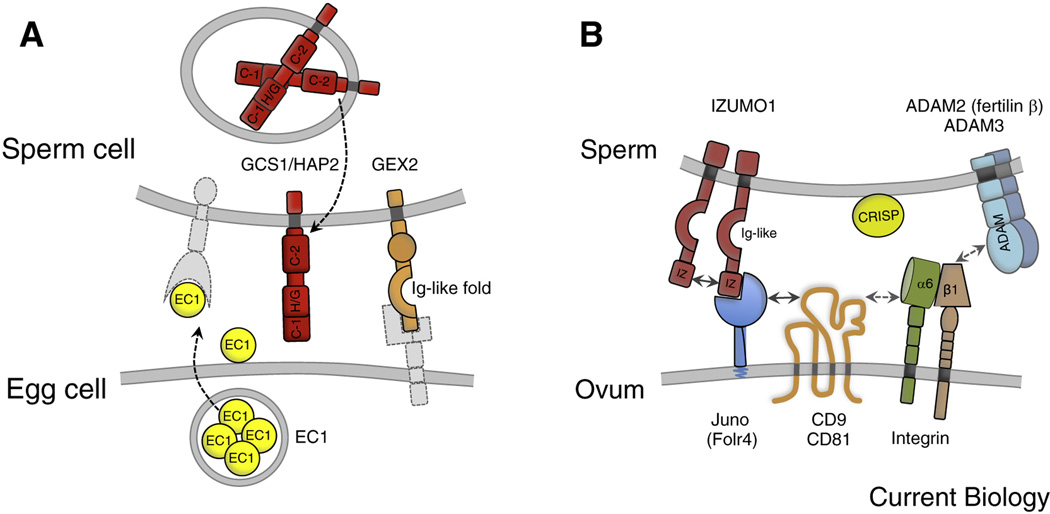
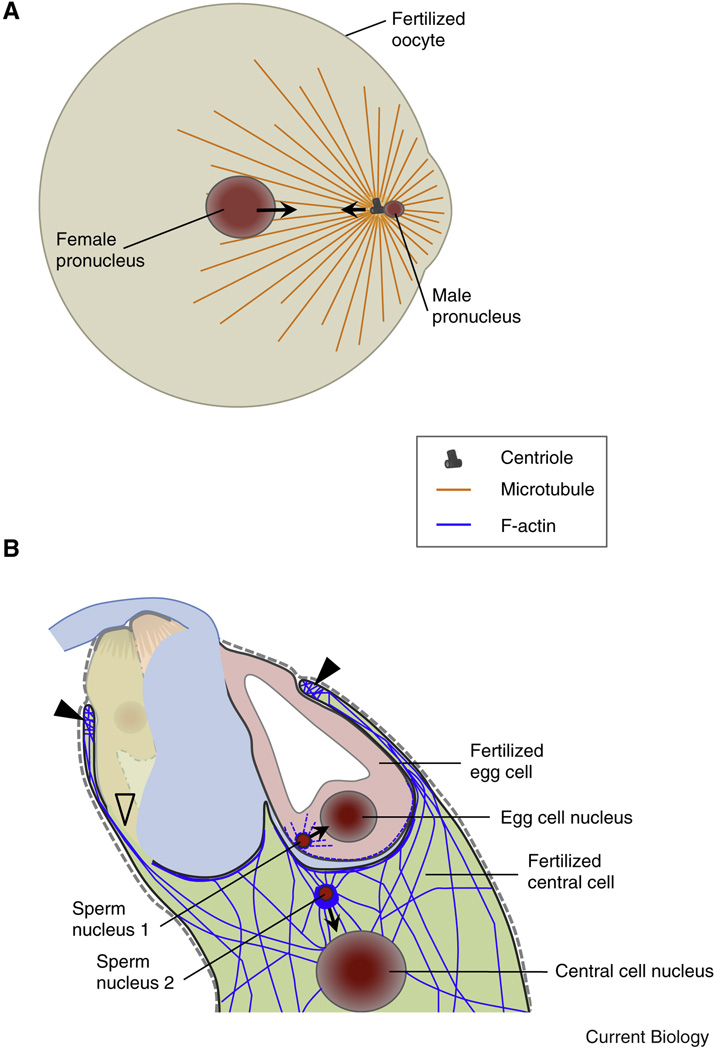
Compared with the animal kingdom, fertilization is particularly complex in flowering plants (angiosperms). Sperm cells of angiosperms have lost their motility and require transportation as a passive cargo by the pollen tube cell to the egg apparatus (egg cell and accessory synergid cells). Sperm cell release from the pollen tube occurs after intensive communication between the pollen tube cell and the receptive synergid, culminating in the lysis of both interaction partners. Following release of the two sperm cells, they interact and fuse with two dimorphic female gametes (the egg and the central cell) forming the major seed components embryo and endosperm, respectively. This process is known as double fertilization. Here, we review the current understanding of the processes of sperm cell reception, gamete interaction, their pre-fertilization activation and fusion, as well as the mechanisms plants use to prevent the fusion of egg cells with multiple sperm cells. The role of Ca(2+) is highlighted in these various processes and comparisons are drawn between fertilization mechanisms in flowering plants and other eukaryotes, including mammals.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Twell D. Male gametogenesis and germline specification in flowering plants. Sex. Plant Reprod. 2011;24:149–160. - PubMed
-
- Sprunck S, Gross-Hardt R. Nuclear behavior, cell polarity, and cell specification in the female gametophyte. Sex. Plant Reprod. 2011;24:123–136. - PubMed
-
- Sprunck S, Hackenberg T, Englhart M, Vogler F. Same same but different: sperm-activating EC1 and ECA1 gametogenesis-related family proteins. Biochem. Soc. Trans. 2014;42:401–407. - PubMed
-
- Dresselhaus T, Franklin-Tong N. Male-female crosstalk during pollen germination, tube growth and guidance, and double fertilization. Mol. Plant. 2013;6:1018–1036. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
