

Circadian control of oscillations in mitochondrial rate-limiting enzymes and nutrient utilization by PERIOD proteins
- PMID: 26862173
- PMCID: PMC4812734
- DOI: 10.1073/pnas.1519650113
Circadian control of oscillations in mitochondrial rate-limiting enzymes and nutrient utilization by PERIOD proteins
Abstract
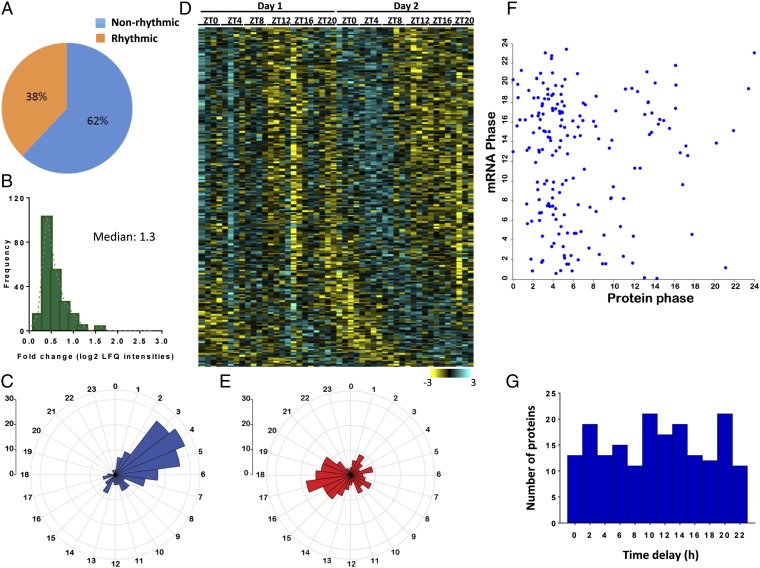
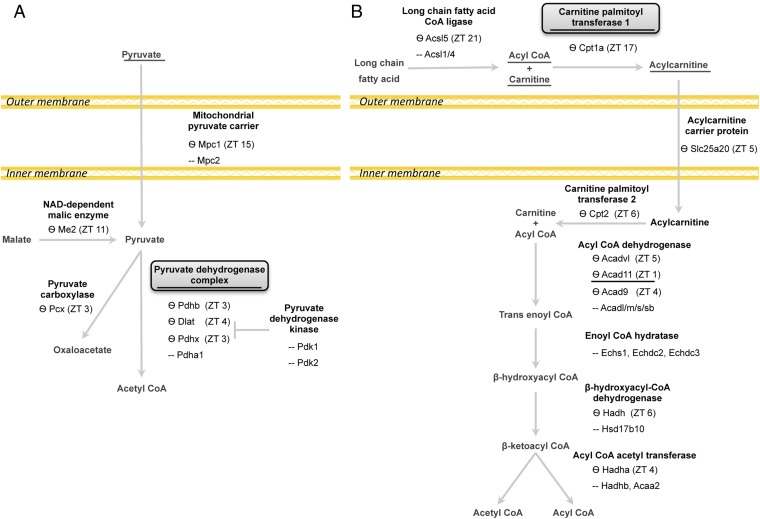
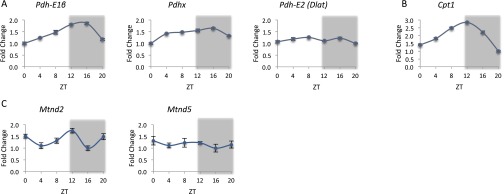
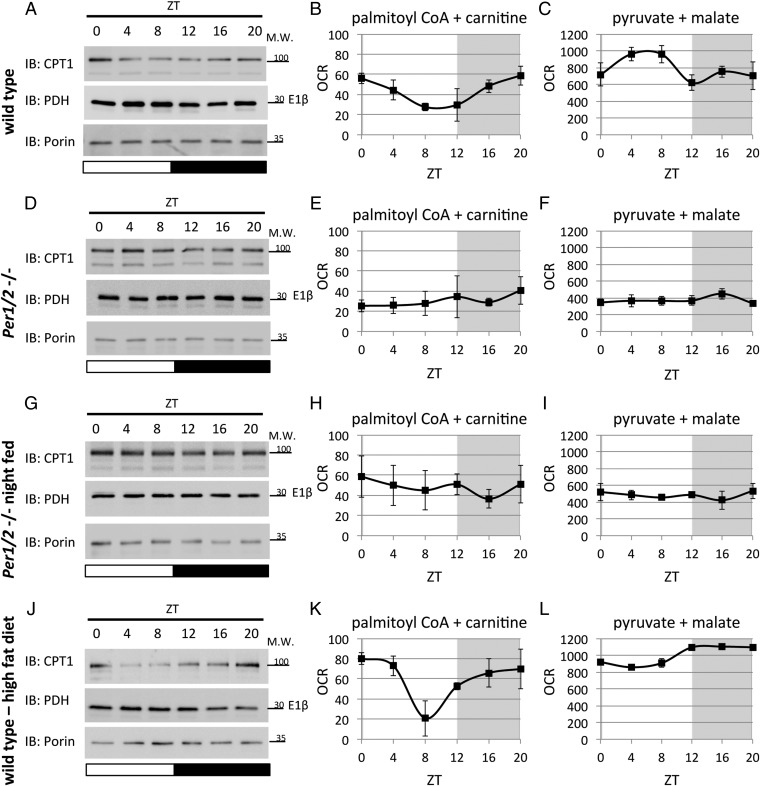
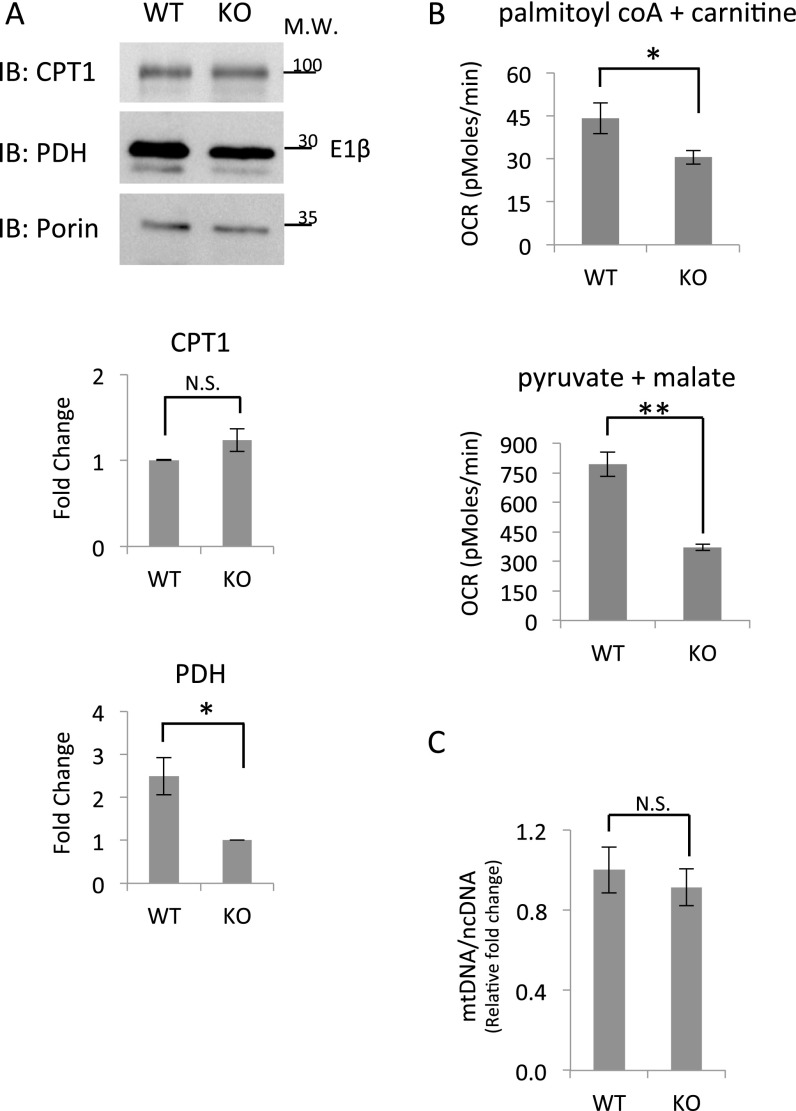
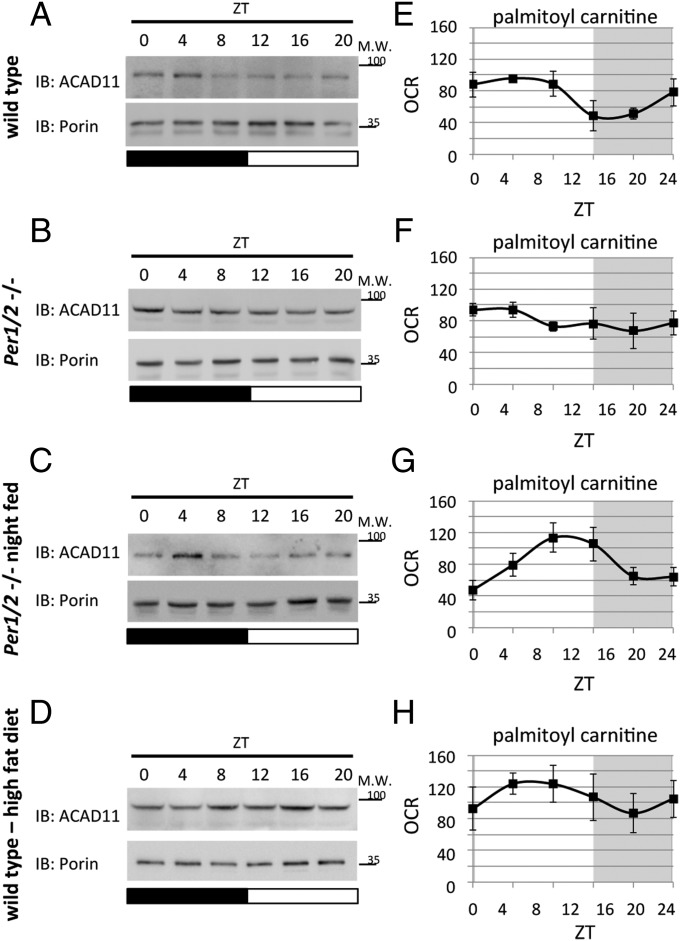
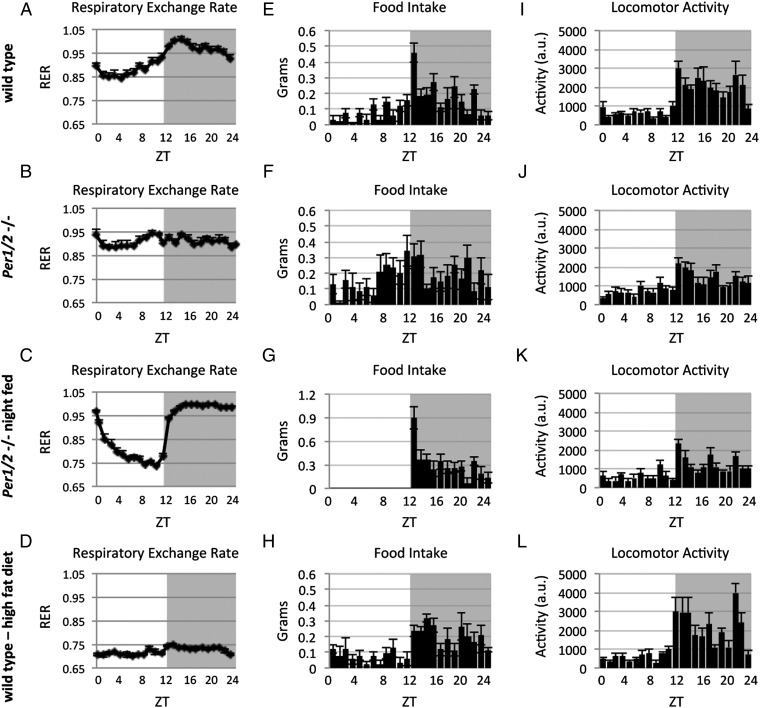
Mitochondria are major suppliers of cellular energy through nutrients oxidation. Little is known about the mechanisms that enable mitochondria to cope with changes in nutrient supply and energy demand that naturally occur throughout the day. To address this question, we applied MS-based quantitative proteomics on isolated mitochondria from mice killed throughout the day and identified extensive oscillations in the mitochondrial proteome. Remarkably, the majority of cycling mitochondrial proteins peaked during the early light phase. We found that rate-limiting mitochondrial enzymes that process lipids and carbohydrates accumulate in a diurnal manner and are dependent on the clock proteins PER1/2. In this conjuncture, we uncovered daily oscillations in mitochondrial respiration that peak during different times of the day in response to different nutrients. Notably, the diurnal regulation of mitochondrial respiration was blunted in mice lacking PER1/2 or on a high-fat diet. We propose that PERIOD proteins optimize mitochondrial metabolism to daily changes in energy supply/demand and thereby, serve as a rheostat for mitochondrial nutrient utilization.
Keywords: PERIOD proteins; circadian rhythm; metabolism; mitochondria; proteomics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Comment in
-
Circadian clock, nutrient quality, and eating pattern tune diurnal rhythms in the mitochondrial proteome.Proc Natl Acad Sci U S A. 2016 Mar 22;113(12):3127-9. doi: 10.1073/pnas.1601786113. Epub 2016 Mar 15. Proc Natl Acad Sci U S A. 2016. PMID: 26979954 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
