

Specificity and disease in the ubiquitin system
- PMID: 26862208
- PMCID: PMC5264512
- DOI: 10.1042/BST20150209
Specificity and disease in the ubiquitin system
Abstract
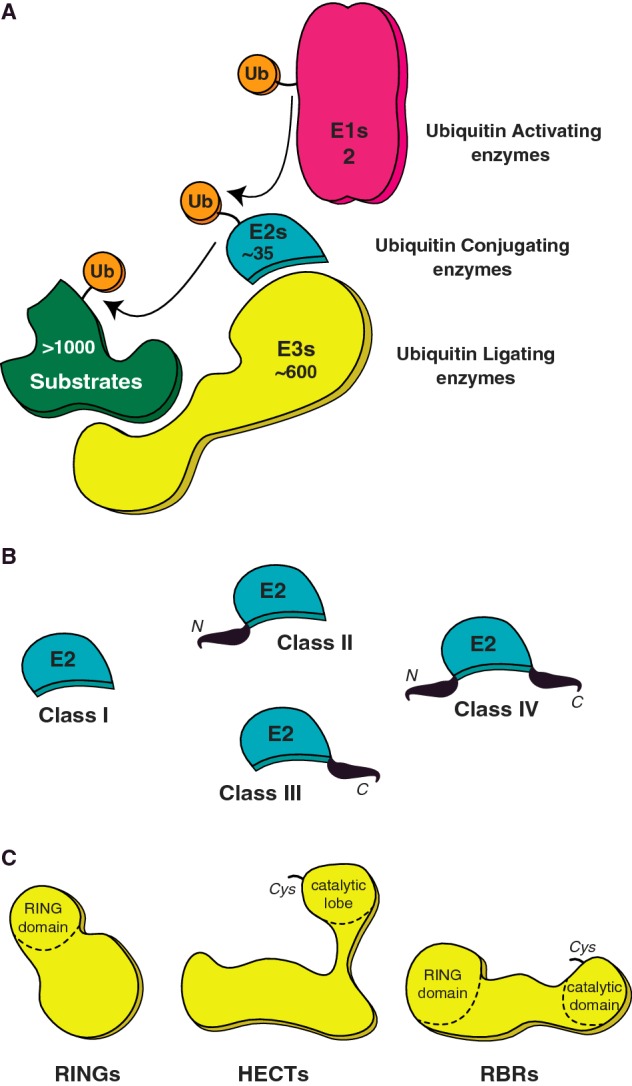
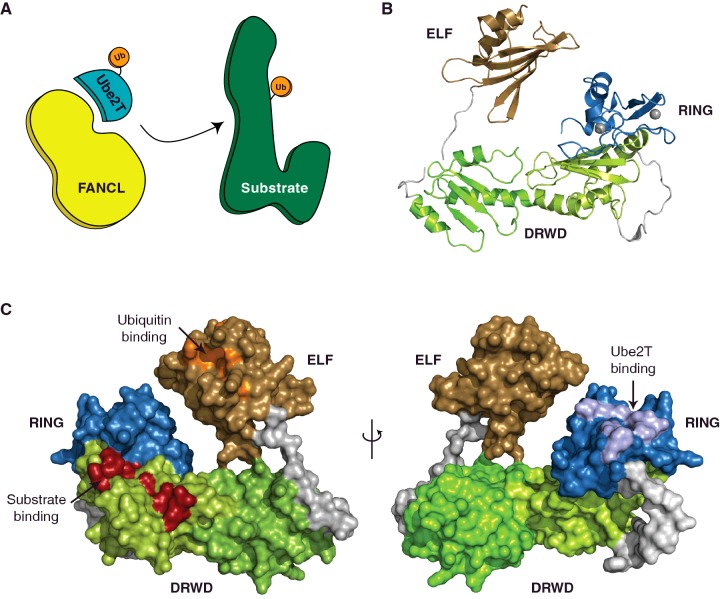
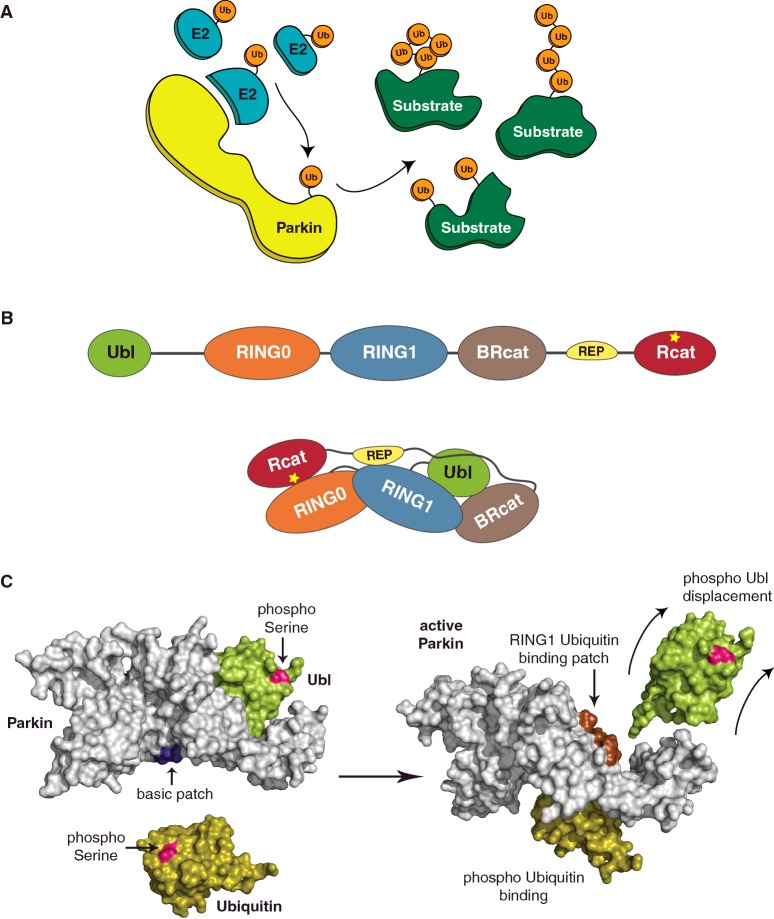
Post-translational modification (PTM) of proteins by ubiquitination is an essential cellular regulatory process. Such regulation drives the cell cycle and cell division, signalling and secretory pathways, DNA replication and repair processes and protein quality control and degradation pathways. A huge range of ubiquitin signals can be generated depending on the specificity and catalytic activity of the enzymes required for attachment of ubiquitin to a given target. As a consequence of its importance to eukaryotic life, dysfunction in the ubiquitin system leads to many disease states, including cancers and neurodegeneration. This review takes a retrospective look at our progress in understanding the molecular mechanisms that govern the specificity of ubiquitin conjugation.
Keywords: E3 ligase; Fanconi anemia; Parkin; RBR; RING; ubiquitin.
© 2016 Authors; published by Portland Press Limited.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials