

doi: 10.1128/JVI.02832-15.
Print 2016 May.
Hepatitis B Virus Infection of a Mouse Hepatic Cell Line Reconstituted with Human Sodium Taurocholate Cotransporting Polypeptide
Affiliations
- PMID: 26865711
- PMCID: PMC4836309
- DOI: 10.1128/JVI.02832-15
Item in Clipboard
Hepatitis B Virus Infection of a Mouse Hepatic Cell Line Reconstituted with Human Sodium Taurocholate Cotransporting Polypeptide
J Virol.
.
Abstract
Hepatitis B virus (HBV) enters hepatocytes via its receptor, human sodium taurocholate cotransporting polypeptide (hNTCP). So far, HBV infection has been achieved only in human hepatic cells reconstituted with hNTCP and not in cells of mouse origin. Here, the first mouse liver cell line (AML12) which gains susceptibility to HBV upon hNTCP expression is described. Thus, HBV infection of receptor-expressing mouse hepatocytes does not principally require a human cofactor but can be triggered by endogenous murine determinants.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
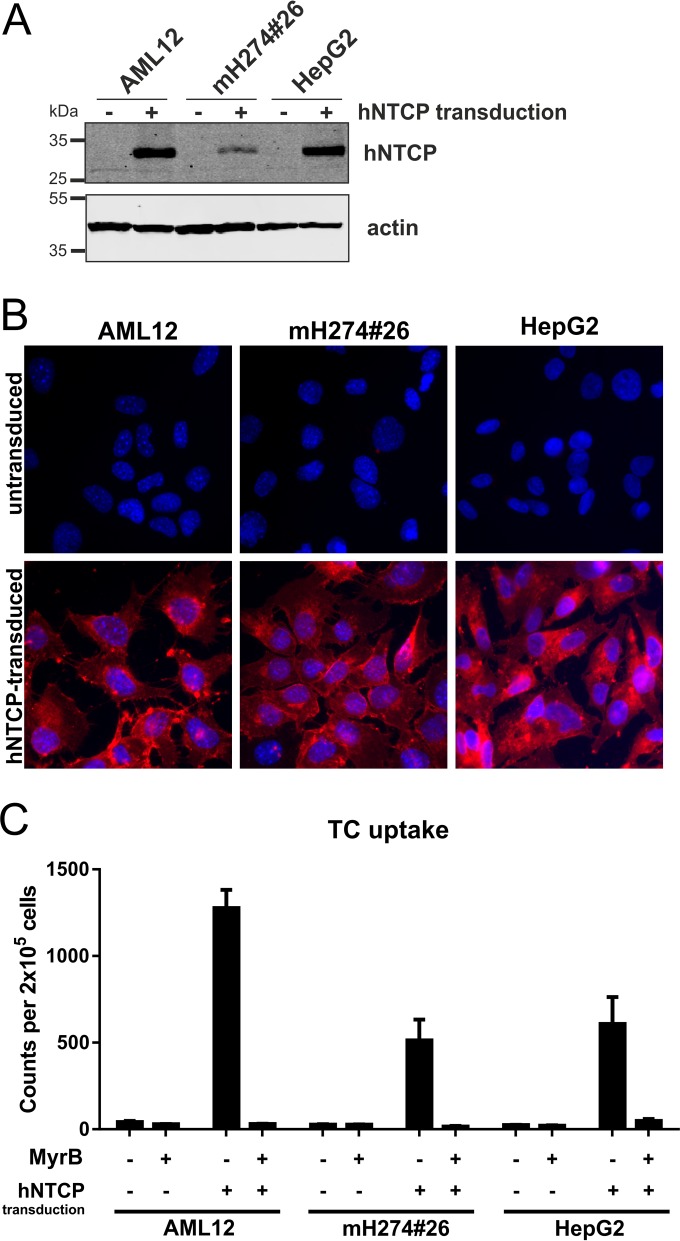
Characterization of mouse and human cell lines stably expressing hNTCP. AML12 (obtained from ATCC CRL-2254), mH274#26 (8), and HepG2 cells were transduced with lentiviruses coexpressing hNTCP and a puromycin resistance gene and selected to establish a stable cell pool. (A) Total cell lysates of the transduced (+) or parental (−) cells were treated with PNGase F (New England BioLabs) and analyzed by Western blotting using a hNTCP-specific antibody (Sigma-Aldrich). (B) The respective transduced (lower row) and parental (upper row) cell lines were incubated for 30 min with 200 µM atto565-labeled Myrcludex B, washed, and analyzed by fluorescence microscopy. (C) Cell lines were incubated with 3H-TC (Hartmann Analytic) for 15 min at 37°C, washed, and lysed, and scintillation counts of the total lysates were measured and the results normalized to the cell number. TC uptake was inhibited by treatment of the cells with 2 μM MyrB (13).
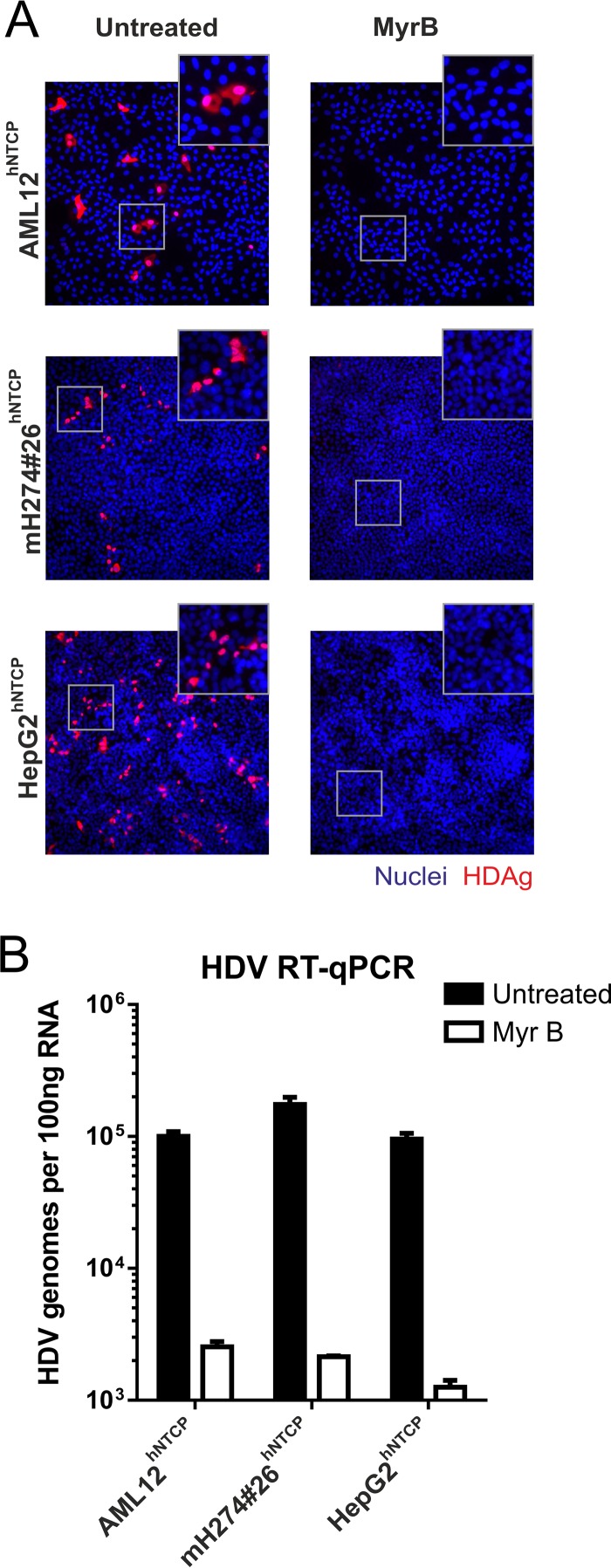
HDV infection of stable hNTCP-expressing mouse and human liver cell lines. AML12hNTCP, mH274#26hNTCP, and HepG2hNTCP cells were seeded in 24-well plates and infected 2 days later with HDV by inoculation of the cells with 4 IU/cell HDV (8) in the presence of 4% polyethylene glycol (PEG) in medium supplemented with 2% dimethyl sulfoxide (DMSO) for 16 h. Entry inhibition was achieved with 500 nM Myrcludex B 30 min prior and during virus inoculation. (A) At day 5 postinfection, cells were fixed with 4% paraformaldehyde (PFA) and permeabilized and HDAg was stained using patient serum (Vuda) and a fluorescently labeled goat-anti-human secondary antibody (Thermo Fisher Scientific). Images (×200 magnification) were acquired on an inverted fluorescence microscope (Leica). (B) Total RNA was extracted from the cells at day 5 postinfection, reverse transcribed, and analyzed by quantitative PCR using primers and probes specific for HDV as previously described (8, 18).
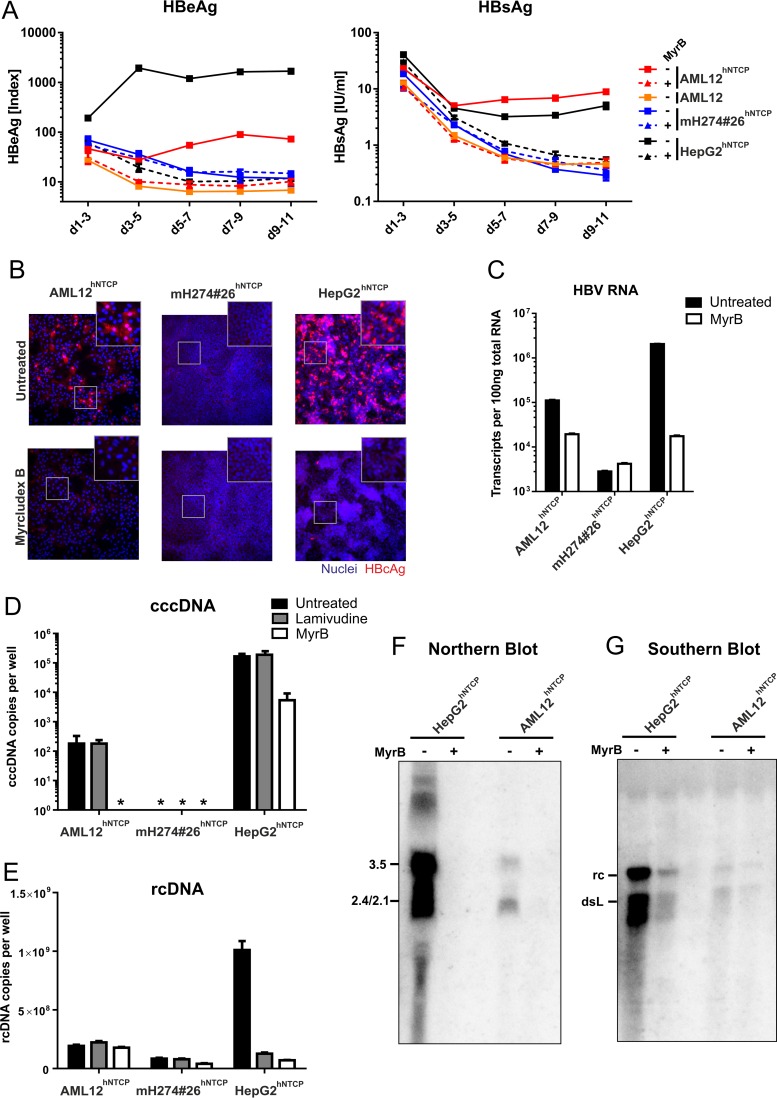
HBV infection of stable hNTCP-expressing cell lines. AML12, AML12hNTCP, mH274#26hNTCP, and HepG2hNTCP cells were seeded in 24-well plates and infected 2 days later by inoculation with HBV at a multiplicity of genome equivalents (MGE) of 1,500 (genotype D, cell culture derived) in the presence of 4% PEG and 2% DMSO for 16 h as described previously (8). For entry inhibition, cells were treated with 500 nM Myrcludex B 30 min prior and during virus inoculation. (A) Supernatants of the cells were collected every second day, and secreted viral markers HBeAg (left) and HBsAg (right) were quantified by ELISA as previously described (8). Each data point represents the mean of the results of three biological replicates. d, day. (B) At day 11 postinfection, cells were fixed and immunostained with an antibody (Dako) recognizing viral HBcAg (red, ×200 magnification). (C) RNA from infected cells was extracted, reverse transcribed, and quantified using primers binding to the HBV core open reading frame (ORF), thereby detecting pregenomic, precore, and possibly spliced HBV RNA (4). (D and E) Total DNA was extracted from cells at day 10 postinfection. Lamivudine (10 μM) was added at day 3 postinfection. rcDNA was quantified with HBV-specific primers (E). Following digestion of rcDNA with T5 exonuclease (New England BioLabs), cccDNA (D) was specifically quantified using primers spanning the gap region (Bingqian Qu, unpublished data). Values below the limit of quantitation are marked by asterisks (*). (F and G) AML12hNTCP and HepG2hNTCP cells were seeded in 10 cm-diameter dishes and infected with HBV at an MGE of 1,500 in the presence or absence of 1 μM MyrB as described above. (F) Eight days after infection, total cellular RNA was extracted, separated on a 1.2% agarose gel, blotted on a nylon membrane, and analyzed with a 32P-labeled HBV-specific probe of genotype D (Northern blotting). (G) Ten days after infection, cytosolic DNA was extracted by treatment of the cells with NP-40 lysis buffer, centrifugation, proteinase K treatment of the supernatant, and subsequent phenol-chloroform DNA extraction. The DNA was separated on a 1.2% agarose gel and analyzed by Southern blotting as described above.

HBV infection of single-cell clones derived from AML12hNTCP. Single-cell clones of AML12hNTCP were picked, expanded, seeded in 24-well plates and analyzed for their ability to transport 3H-TC (see Fig. 1 for details) (A) or infected by inoculation with HBV at an MGE of 1,500 in the presence or absence of MyrB as described in the Fig. 3 legend (B, C, and D). The supernatant of the infected cells was collected from day 7 to day 9 postinfection and analyzed for secreted HBeAg (B) and HBsAg (C). Cells were fixed and stained with an antibody against viral HBcAg (D).
References
-
- Ni Y, Lempp FA, Mehrle S, Nkongolo S, Kaufman C, Falth M, Stindt J, Koniger C, Nassal M, Kubitz R, Sultmann H, Urban S. 2014. Hepatitis B and D viruses exploit sodium taurocholate co-transporting polypeptide for species-specific entry into hepatocytes. Gastroenterology 146:1070–1083. doi: 10.1053/j.gastro.2013.12.024. - DOI - PubMed
-
- Yan H, Zhong G, Xu G, He W, Jing Z, Gao Z, Huang Y, Qi Y, Peng B, Wang H, Fu L, Song M, Chen P, Gao W, Ren B, Sun Y, Cai T, Feng X, Sui J, Li W. 2012. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. Elife 1:e00049. doi: 10.7554/eLife.00049. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
