

Microtubule and Actin Interplay Drive Intracellular c-Src Trafficking
- PMID: 26866809
- PMCID: PMC4750819
- DOI: 10.1371/journal.pone.0148996
Microtubule and Actin Interplay Drive Intracellular c-Src Trafficking
Abstract
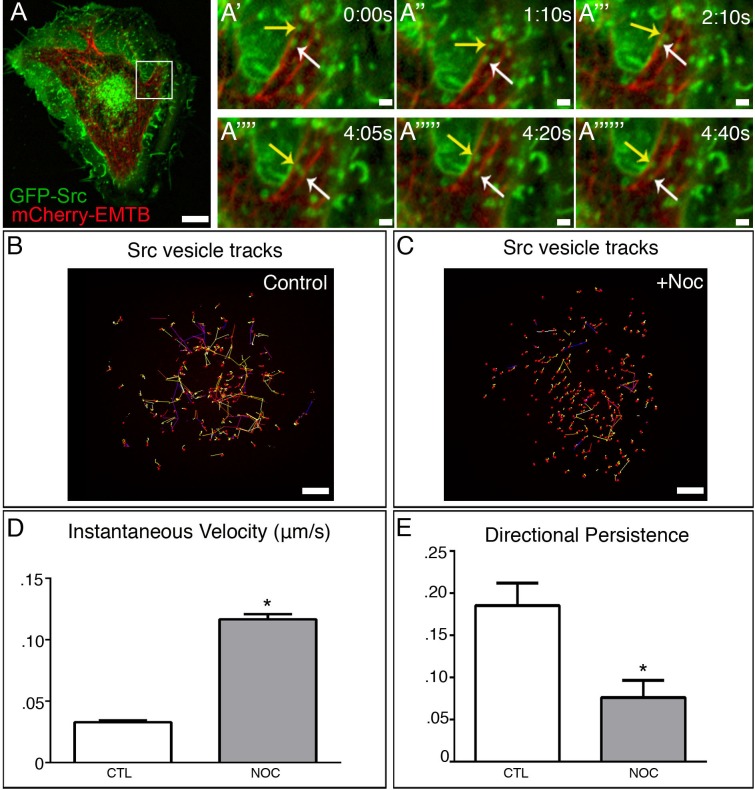
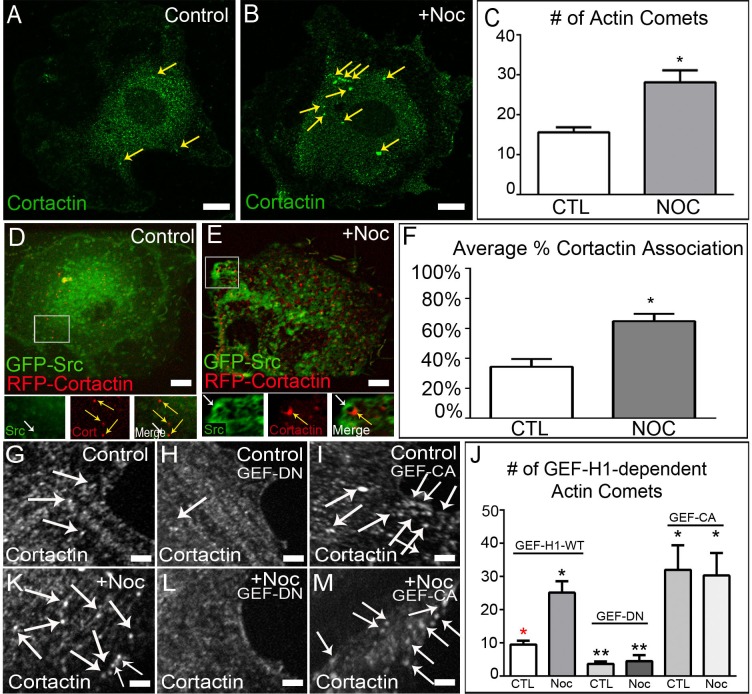
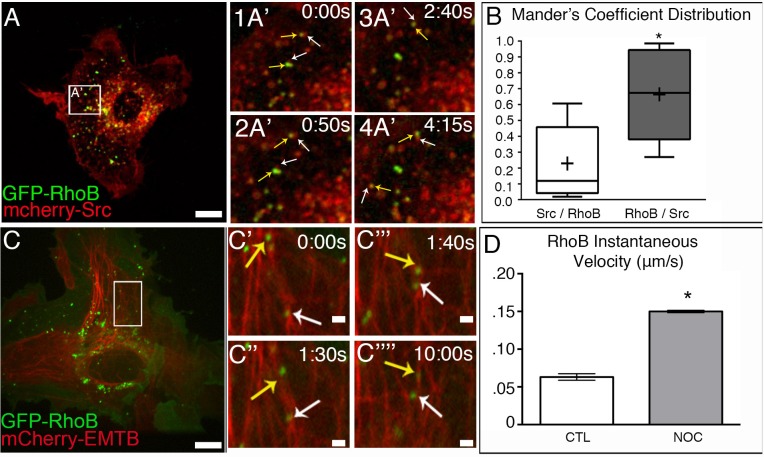
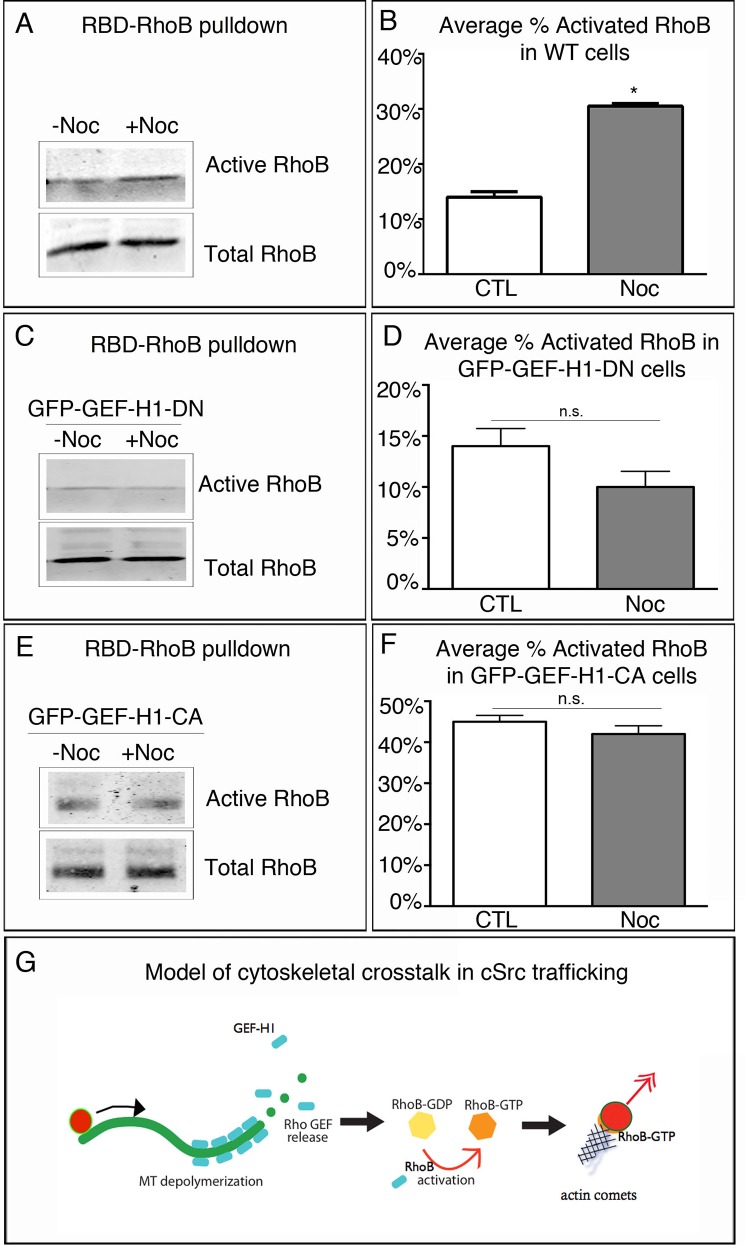
The proto-oncogene c-Src is involved in a variety of signaling processes. Therefore, c-Src spatiotemporal localization is critical for interaction with downstream targets. However, the mechanisms regulating this localization have remained elusive. Previous studies have shown that c-Src trafficking is a microtubule-dependent process that facilitates c-Src turnover in neuronal growth cones. As such, microtubule depolymerization lead to the inhibition of c-Src recycling. Alternatively, c-Src trafficking was also shown to be regulated by RhoB-dependent actin polymerization. Our results show that c-Src vesicles primarily exhibit microtubule-dependent trafficking; however, microtubule depolymerization does not inhibit vesicle movement. Instead, vesicular movement becomes both faster and less directional. This movement was associated with actin polymerization directly at c-Src vesicle membranes. Interestingly, it has been shown previously that c-Src delivery is an actin polymerization-dependent process that relies on small GTPase RhoB at c-Src vesicles. In agreement with this finding, microtubule depolymerization induced significant activation of RhoB, together with actin comet tail formation. These effects occurred downstream of GTP-exchange factor, GEF-H1, which was released from depolymerizing MTs. Accordingly, GEF-H1 activity was necessary for actin comet tail formation at the Src vesicles. Our results indicate that regulation of c-Src trafficking requires both microtubules and actin polymerization, and that GEF-H1 coordinates c-Src trafficking, acting as a molecular switch between these two mechanisms.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
