

Induction of virulence factors in Giardia duodenalis independent of host attachment
- PMID: 26867958
- PMCID: PMC4751611
- DOI: 10.1038/srep20765
Induction of virulence factors in Giardia duodenalis independent of host attachment
Abstract
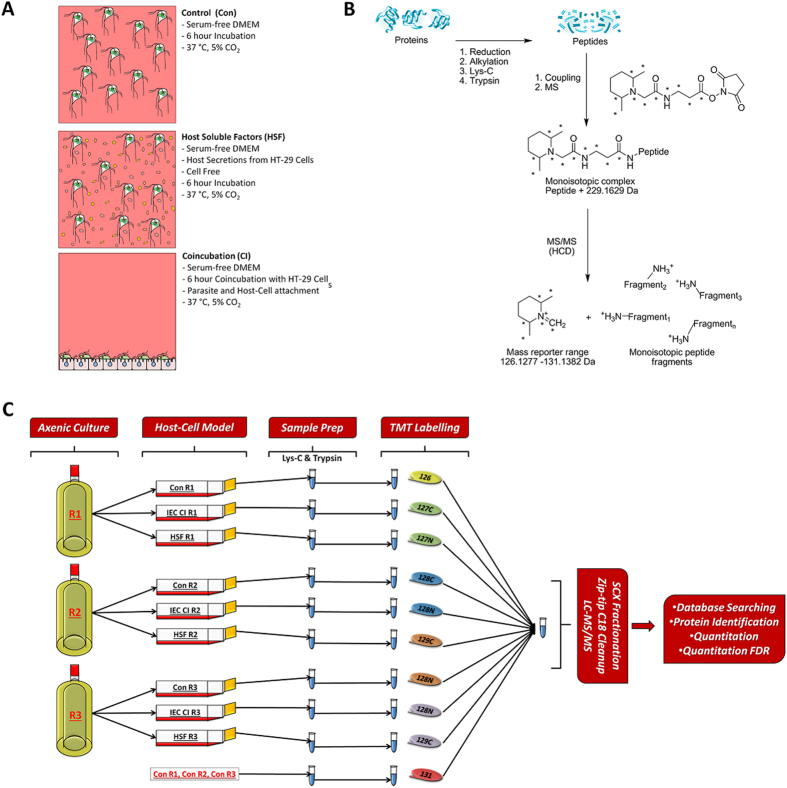
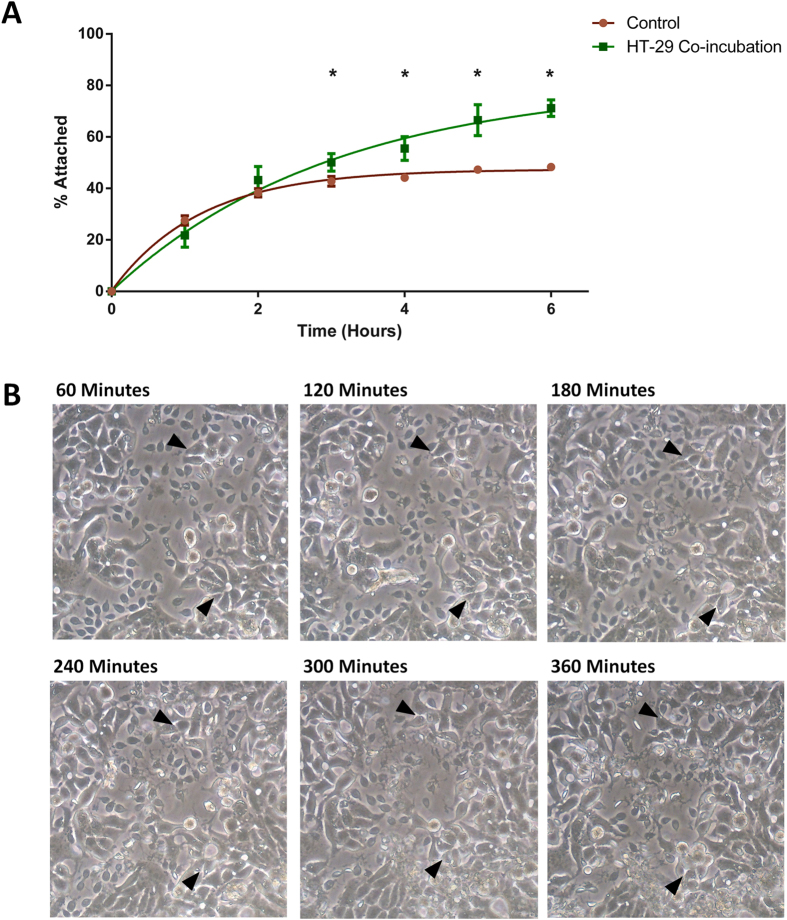
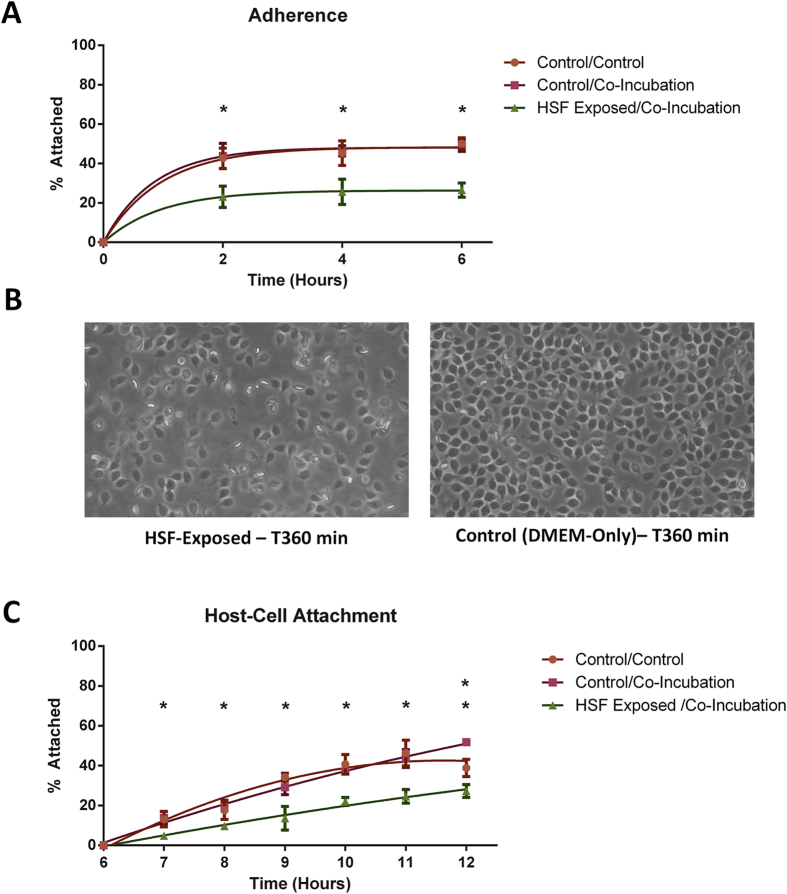
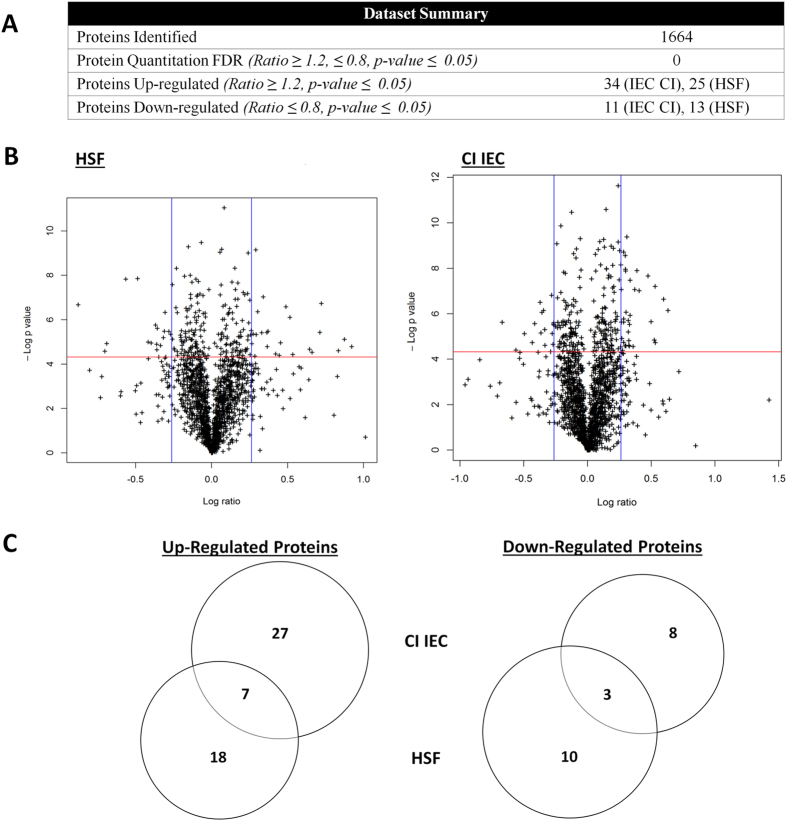
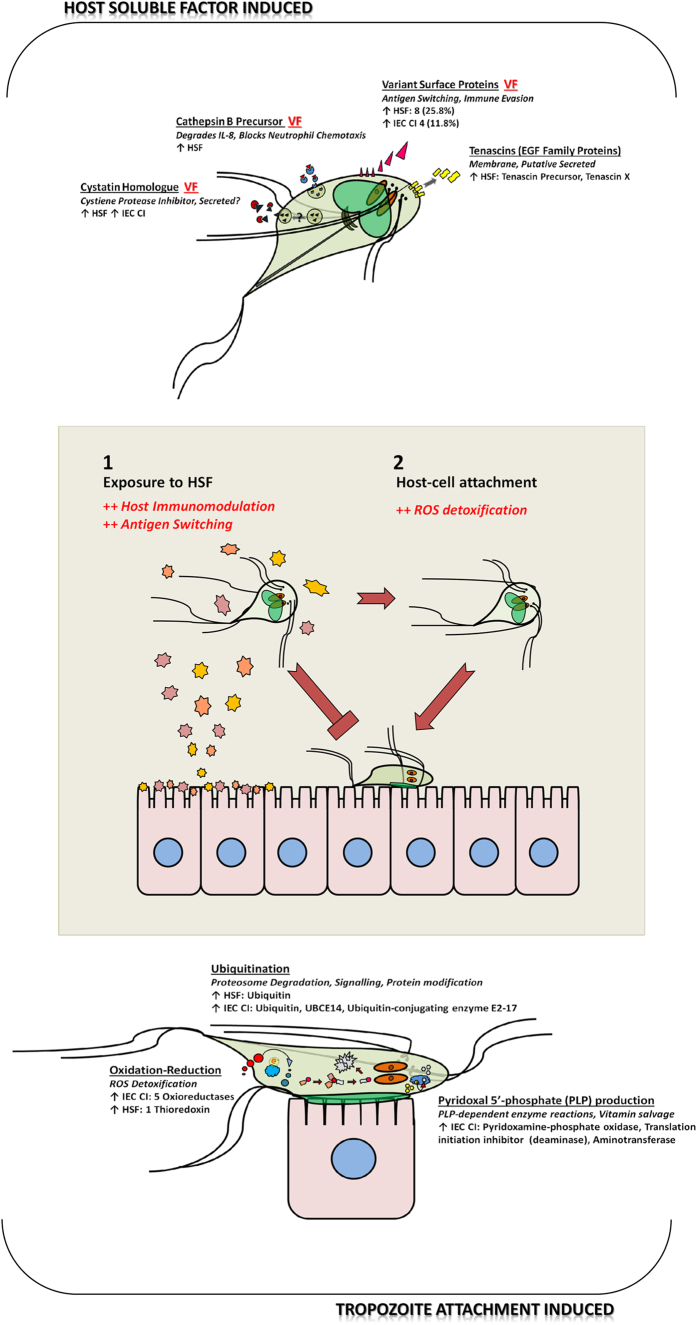
Giardia duodenalis is responsible for the majority of parasitic gastroenteritis in humans worldwide. Host-parasite interaction models in vitro provide insights into disease and virulence and help us to understand pathogenesis. Using HT-29 intestinal epithelial cells (IEC) as a model we have demonstrated that initial sensitisation by host secretions reduces proclivity for trophozoite attachment, while inducing virulence factors. Host soluble factors triggered up-regulation of membrane and secreted proteins, including Tenascins, Cathepsin-B precursor, cystatin, and numerous Variant-specific Surface Proteins (VSPs). By comparison, host-cell attached trophozoites up-regulated intracellular pathways for ubiquitination, reactive oxygen species (ROS) detoxification and production of pyridoxal phosphate (PLP). We reason that these results demonstrate early pathogenesis in Giardia involves two independent host-parasite interactions. Motile trophozoites respond to soluble secreted signals, which deter attachment and induce expression of virulence factors. Trophozoites attached to host cells, in contrast, respond by up-regulating intracellular pathways involved in clearance of ROS, thus anticipating the host defence response.
Figures
Similar articles
-
Expression and secretion of the Giardia duodenalis variant surface protein 9B10A by transfected trophozoites causes damage to epithelial cell monolayers mediated by protease activity.Exp Parasitol. 2017 Aug;179:49-64. doi: 10.1016/j.exppara.2017.06.006. Epub 2017 Jun 28. Exp Parasitol. 2017. PMID: 28668253
-
Genotyping and Descriptive Proteomics of a Potential Zoonotic Canine Strain of Giardia duodenalis, Infective to Mice.PLoS One. 2016 Oct 19;11(10):e0164946. doi: 10.1371/journal.pone.0164946. eCollection 2016. PLoS One. 2016. PMID: 27760188 Free PMC article.
-
Giardia duodenalis enolase is secreted as monomer during trophozoite-epithelial cell interactions, activates plasminogen and induces necroptotic damage.Front Cell Infect Microbiol. 2022 Aug 25;12:928687. doi: 10.3389/fcimb.2022.928687. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 36093180 Free PMC article.
-
Giardia duodenalis: Role of secreted molecules as virulent factors in the cytotoxic effect on epithelial cells.Adv Parasitol. 2019;106:129-169. doi: 10.1016/bs.apar.2019.07.003. Epub 2019 Aug 20. Adv Parasitol. 2019. PMID: 31630757 Review.
-
Giardia's Epithelial Cell Interaction In Vitro: Mimicking Asymptomatic Infection?Front Cell Infect Microbiol. 2017 Sep 26;7:421. doi: 10.3389/fcimb.2017.00421. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 29018775 Free PMC article. Review.
Cited by
-
Host-Parasite Interaction of Atlantic salmon (Salmo salar) and the Ectoparasite Neoparamoeba perurans in Amoebic Gill Disease.Front Immunol. 2021 May 31;12:672700. doi: 10.3389/fimmu.2021.672700. eCollection 2021. Front Immunol. 2021. PMID: 34135900 Free PMC article.
-
Helicobacter pylori Pathogenicity Islands and Giardia lamblia Cysteine Proteases in Role of Coinfection and Pathogenesis.Infect Drug Resist. 2022 Jan 6;15:21-34. doi: 10.2147/IDR.S346705. eCollection 2022. Infect Drug Resist. 2022. PMID: 35023934 Free PMC article. Review.
-
Differential protein expression and post-translational modifications in metronidazole-resistant Giardia duodenalis.Gigascience. 2018 Apr 1;7(4):giy024. doi: 10.1093/gigascience/giy024. Gigascience. 2018. PMID: 29688452 Free PMC article.
-
Secreted Giardia intestinalis cysteine proteases disrupt intestinal epithelial cell junctional complexes and degrade chemokines.Virulence. 2018 Dec 31;9(1):879-894. doi: 10.1080/21505594.2018.1451284. Virulence. 2018. PMID: 29726306 Free PMC article.
-
Retortamonads from vertebrate hosts share features of anaerobic metabolism and pre-adaptations to parasitism with diplomonads.Parasitol Int. 2021 Jun;82:102308. doi: 10.1016/j.parint.2021.102308. Epub 2021 Feb 21. Parasitol Int. 2021. PMID: 33626397 Free PMC article.
References
-
- Cotton J. A., Beatty J. K. & Buret A. G. Host parasite interactions and pathophysiology in Giardia infections. Int J Parasitol 41, 925–933 (2011). - PubMed
-
- Buret A. G. Pathophysiology of enteric infections with Giardia duodenalis. Parasite 15, 261–265 (2008). - PubMed
-
- Ringqvist E., Avesson L., Soderbom F. & Svard S. G. Transcriptional changes in Giardia during host-parasite interactions. Int J Parasitol 41, 277–285 (2011). - PubMed
-
- Ferella M. et al. Gene expression changes during Giardia-host cell interactions in serum-free medium. Mol Biochem Parasitol 197, 21–23 (2014). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
