

The chloride-channel blocker 9-anthracenecarboxylic acid reduces the nonlinear capacitance of prestin-associated charge movement
- PMID: 26869218
- PMCID: PMC5111741
- DOI: 10.1111/ejn.13209
The chloride-channel blocker 9-anthracenecarboxylic acid reduces the nonlinear capacitance of prestin-associated charge movement
Abstract
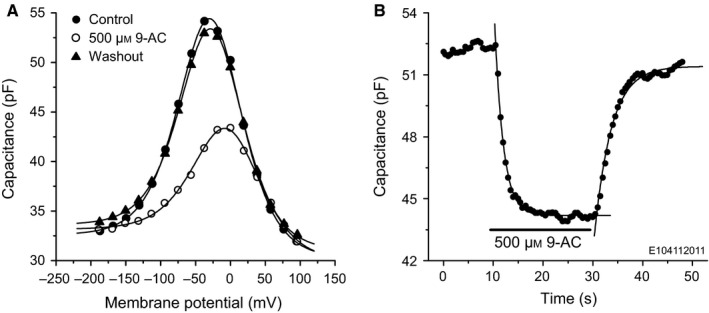
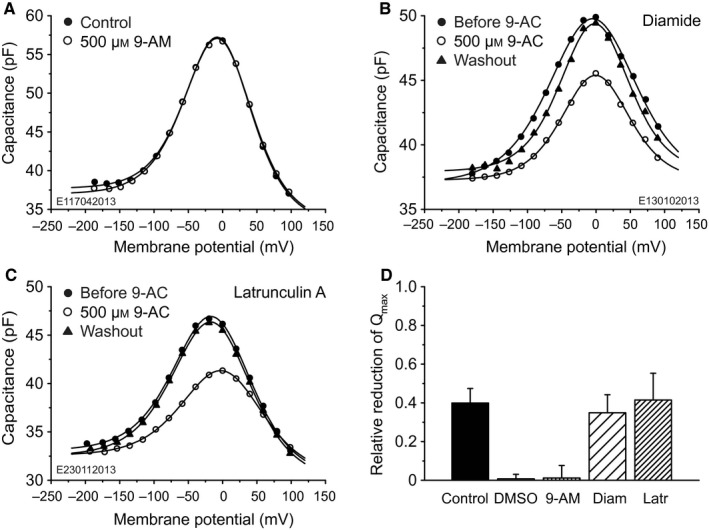
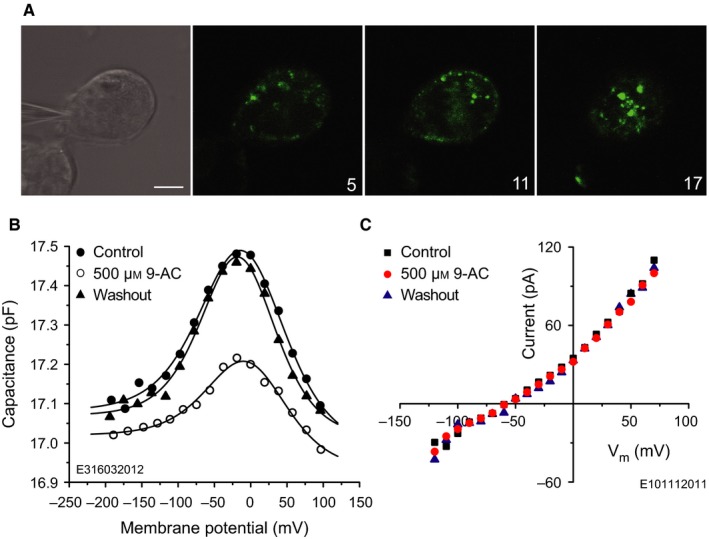
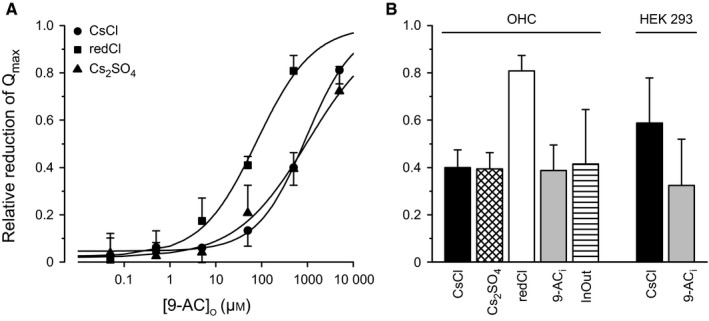
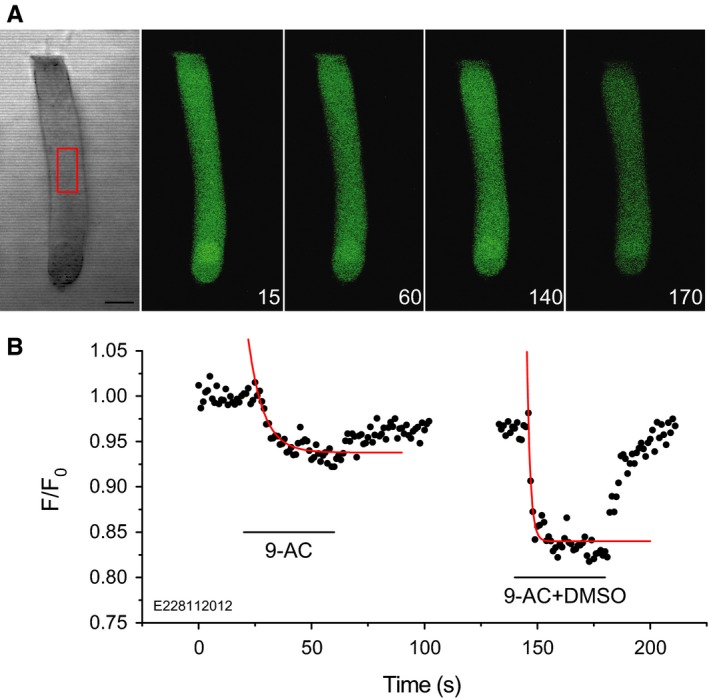
The basis of the extraordinary sensitivity and frequency selectivity of the cochlea is a chloride-sensitive protein called prestin which can produce an electromechanical response and which resides in the basolateral plasma membrane of outer hair cells (OHCs). The compound 9-anthracenecarboxylic acid (9-AC), an inhibitor of chloride channels, has been found to reduce the electromechanical response of the cochlea and the OHC mechanical impedance. To elucidate these 9-AC effects, the functional electromechanical status of prestin was assayed by measuring the nonlinear capacitance of OHCs from the guinea-pig cochlea and of prestin-transfected human embryonic kidney 293 (HEK 293) cells. Extracellular application of 9-AC caused reversible, dose-dependent and chloride-sensitive reduction in OHC nonlinear charge transfer, Qmax . Prestin-transfected cells also showed reversible reduction in Qmax . For OHCs, intracellular 9-AC application as well as reduced intracellular pH had no detectable effect on the reduction in Qmax by extracellularly applied 9-AC. In the prestin-transfected cells, cytosolic application of 9-AC approximately halved the blocking efficacy of extracellularly applied 9-AC. OHC inside-out patches presented the whole-cell blocking characteristics. Disruption of the cytoskeleton by preventing actin polymerization with latrunculin A or by decoupling of spectrin from actin with diamide did not affect the 9-AC-evoked reduction in Qmax . We conclude that 9-AC acts on the electromechanical transducer principally by interaction with prestin rather than acting via the cytoskeleton, chloride channels or pH. The 9-AC block presents characteristics in common with salicylate, but is almost an order of magnitude faster. 9-AC provides a new tool for elucidating the molecular dynamics of prestin function.
Keywords: auditory system; channelopathies; cochlea; electromechanical transduction; sensory hair cells.
© 2016 Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures
Similar articles
-
Chloride Anions Regulate Kinetics but Not Voltage-Sensor Qmax of the Solute Carrier SLC26a5.Biophys J. 2016 Jun 7;110(11):2551-2561. doi: 10.1016/j.bpj.2016.05.002. Biophys J. 2016. PMID: 27276272 Free PMC article.
-
The ultrastructural distribution of prestin in outer hair cells: a post-embedding immunogold investigation of low-frequency and high-frequency regions of the rat cochlea.Eur J Neurosci. 2010 May;31(9):1595-605. doi: 10.1111/j.1460-9568.2010.07182.x. Eur J Neurosci. 2010. PMID: 20525072 Free PMC article.
-
Functional prestin transduction of immature outer hair cells from normal and prestin-null mice.J Assoc Res Otolaryngol. 2008 Sep;9(3):307-20. doi: 10.1007/s10162-008-0121-3. Epub 2008 May 28. J Assoc Res Otolaryngol. 2008. PMID: 18506528 Free PMC article.
-
Control of mammalian cochlear amplification by chloride anions.J Neurosci. 2006 Apr 12;26(15):3992-8. doi: 10.1523/JNEUROSCI.4548-05.2006. J Neurosci. 2006. PMID: 16611815 Free PMC article.
-
Prestin and electromotility may serve multiple roles in cochlear outer hair cells.Hear Res. 2022 Sep 15;423:108428. doi: 10.1016/j.heares.2021.108428. Epub 2021 Dec 26. Hear Res. 2022. PMID: 34987016 Review.
Cited by
-
Chloride Channel and Inflammation-Mediated Pathogenesis of Osteoarthritis.J Inflamm Res. 2022 Feb 11;15:953-964. doi: 10.2147/JIR.S350432. eCollection 2022. J Inflamm Res. 2022. PMID: 35177922 Free PMC article. Review.
-
Prestin kinetics and corresponding frequency dependence augment during early development of the outer hair cell within the mouse organ of Corti.Sci Rep. 2019 Nov 11;9(1):16460. doi: 10.1038/s41598-019-52965-1. Sci Rep. 2019. PMID: 31712635 Free PMC article.
References
-
- Ashmore, J. (2008) Cochlear outer hair cell motility. Physiol. Rev., 88, 173–210. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
