

Mucosal immunoglobulins at respiratory surfaces mark an ancient association that predates the emergence of tetrapods
- PMID: 26869478
- PMCID: PMC4754351
- DOI: 10.1038/ncomms10728
Mucosal immunoglobulins at respiratory surfaces mark an ancient association that predates the emergence of tetrapods
Abstract
Gas-exchange structures are critical for acquiring oxygen, but they also represent portals for pathogen entry. Local mucosal immunoglobulin responses against pathogens in specialized respiratory organs have only been described in tetrapods. Since fish gills are considered a mucosal surface, we hypothesized that a dedicated mucosal immunoglobulin response would be generated within its mucosa on microbial exposure. Supporting this hypothesis, here we demonstrate that following pathogen exposure, IgT(+) B cells proliferate and generate pathogen-specific IgT within the gills of fish, thus providing the first example of locally induced immunoglobulin in the mucosa of a cold-blooded species. Moreover, we demonstrate that gill microbiota is predominantly coated with IgT, thus providing previously unappreciated evidence that the microbiota present at a respiratory surface of a vertebrate is recognized by a mucosal immunoglobulin. Our findings indicate that respiratory surfaces and mucosal immunoglobulins are part of an ancient association that predates the emergence of tetrapods.
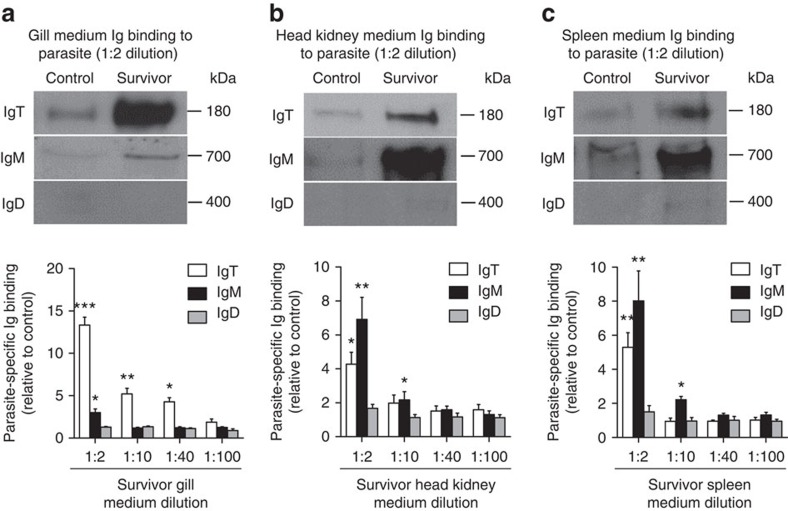
Figures








Similar articles
-
The predominant role of mucosal immunoglobulin IgT in the gills of rainbow trout (Oncorhynchus mykiss) after infection with Flavobacterium columnare.Fish Shellfish Immunol. 2020 Apr;99:654-662. doi: 10.1016/j.fsi.2020.01.044. Epub 2020 Jan 27. Fish Shellfish Immunol. 2020. PMID: 32001351
-
Cellular and humoral factors involved in the response of rainbow trout gills to Ichthyophthirius multifiliis infections: molecular and immunohistochemical studies.Fish Shellfish Immunol. 2011 Mar;30(3):859-69. doi: 10.1016/j.fsi.2011.01.010. Epub 2011 Jan 25. Fish Shellfish Immunol. 2011. PMID: 21272651
-
Teleost skin, an ancient mucosal surface that elicits gut-like immune responses.Proc Natl Acad Sci U S A. 2013 Aug 6;110(32):13097-102. doi: 10.1073/pnas.1304319110. Epub 2013 Jul 24. Proc Natl Acad Sci U S A. 2013. PMID: 23884653 Free PMC article.
-
Immune responses of fish to Ichthyophthirius multifiliis (Ich): A model for understanding immunity against protozoan parasites.Dev Comp Immunol. 2019 Apr;93:93-102. doi: 10.1016/j.dci.2019.01.002. Epub 2019 Jan 7. Dev Comp Immunol. 2019. PMID: 30630003 Review.
-
Immunoglobulins, Mucosal Immunity and Vaccination in Teleost Fish.Front Immunol. 2020 Oct 2;11:567941. doi: 10.3389/fimmu.2020.567941. eCollection 2020. Front Immunol. 2020. PMID: 33123139 Free PMC article. Review.
Cited by
-
Gill and Liver Transcript Expression Changes Associated With Gill Damage in Atlantic Salmon (Salmo salar).Front Immunol. 2022 Mar 28;13:806484. doi: 10.3389/fimmu.2022.806484. eCollection 2022. Front Immunol. 2022. PMID: 35418993 Free PMC article.
-
Why Does Piscirickettsia salmonis Break the Immunological Paradigm in Farmed Salmon? Biological Context to Understand the Relative Control of Piscirickettsiosis.Front Immunol. 2022 Mar 21;13:856896. doi: 10.3389/fimmu.2022.856896. eCollection 2022. Front Immunol. 2022. PMID: 35386699 Free PMC article. Review.
-
Recent Advances on Phagocytic B Cells in Teleost Fish.Front Immunol. 2020 May 27;11:824. doi: 10.3389/fimmu.2020.00824. eCollection 2020. Front Immunol. 2020. PMID: 32536909 Free PMC article. Review.
-
Specific and Efficient Uptake of Surfactant-Free Poly(Lactic Acid) Nanovaccine Vehicles by Mucosal Dendritic Cells in Adult Zebrafish after Bath Immersion.Front Immunol. 2017 Feb 27;8:190. doi: 10.3389/fimmu.2017.00190. eCollection 2017. Front Immunol. 2017. PMID: 28289416 Free PMC article.
-
Comparative transcriptome analysis of the gills of Procambarus clarkii provide novel insights into the response mechanism of ammonia stress tolerance.Mol Biol Rep. 2021 Mar;48(3):2611-2618. doi: 10.1007/s11033-021-06315-y. Epub 2021 Apr 3. Mol Biol Rep. 2021. PMID: 33811573
References
-
- Glover C. N., Bucking C. & Wood C. M. The skin of fish as a transport epithelium: a review. J. Comp. Physiol. B 183, 877–891 (2013). - PubMed
-
- Aquilino C., Castro R., Fischer U. & Tafalla C. Transcriptomic responses in rainbow trout gills upon infection with viral hemorrhagic septicemia virus (VHSV). Dev. Comp. Immunol. 44, 12–20 (2014). - PubMed
-
- Pennacchi Y., Leef M. J., Crosbie P. B., Nowak B. F. & Bridle A. R. Evidence of immune and inflammatory processes in the gills of AGD-affected Atlantic salmon, Salmo salar L. Fish Shellfish Immunol. 36, 563–570 (2014). - PubMed
-
- Valenzuela-Miranda D., Boltana S., Cabrejos M. E., Yanez J. M. & Gallardo-Escarate C. High-throughput transcriptome analysis of ISAV-infected Atlantic salmon Salmo salar unravels divergent immune responses associated to head-kidney, liver and gills tissues. Fish Shellfish Immunol. 45, 367–377 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
