

Creating Order from Chaos: Epigenome Dynamics in Plants with Complex Genomes
- PMID: 26869701
- PMCID: PMC4790878
- DOI: 10.1105/tpc.15.00911
Creating Order from Chaos: Epigenome Dynamics in Plants with Complex Genomes
Abstract
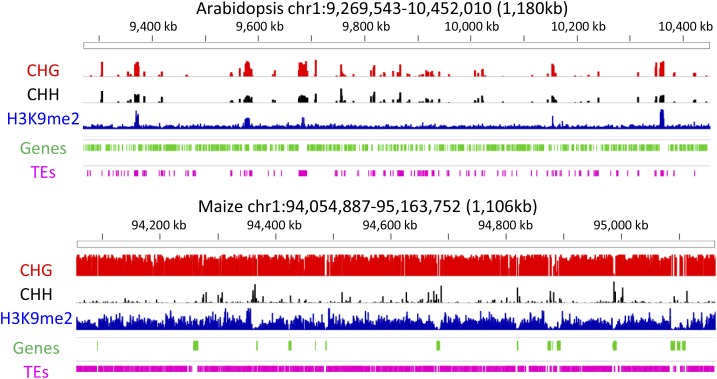
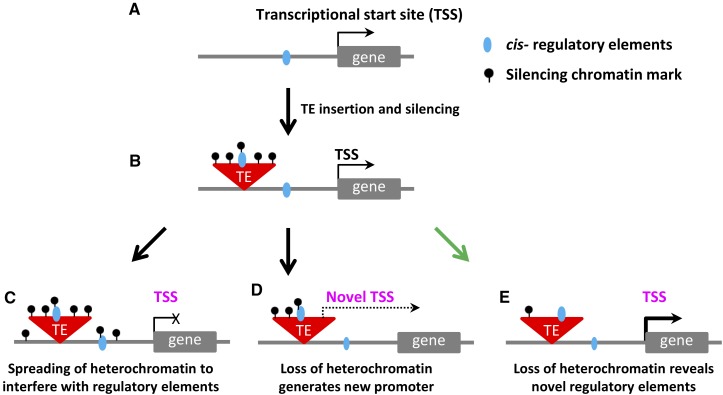
Flowering plants have strikingly distinct genomes, although they contain a similar suite of expressed genes. The diversity of genome structures and organization is largely due to variation in transposable elements (TEs) and whole-genome duplication (WGD) events. We review evidence that chromatin modifications and epigenetic regulation are intimately associated with TEs and likely play a role in mediating the effects of WGDs. We hypothesize that the current structure of a genome is the result of various TE bursts and WGDs and it is likely that the silencing mechanisms and the chromatin structure of a genome have been shaped by these events. This suggests that the specific mechanisms targeting chromatin modifications and epigenomic patterns may vary among different species. Many crop species have likely evolved chromatin-based mechanisms to tolerate silenced TEs near actively expressed genes. These interactions of heterochromatin and euchromatin are likely to have important roles in modulating gene expression and variability within species.
© 2016 American Society of Plant Biologists. All rights reserved.
Figures

Comment in
-
The Plant Cell Reviews Small RNA and Chromatin Dynamics: From Small Genetic Circuits to Complex Genomes.Plant Cell. 2016 Feb;28(2):269-71. doi: 10.1105/tpc.16.00113. Epub 2016 Feb 11. Plant Cell. 2016. PMID: 26869698 Free PMC article. No abstract available.
References
-
- Adams K.L., Wendel J.F. (2005). Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 8: 135–141. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
