

Chronic central serotonin depletion attenuates ventilation and body temperature in young but not adult Tph2 knockout rats
- PMID: 26869713
- PMCID: PMC4855209
- DOI: 10.1152/japplphysiol.01015.2015
Chronic central serotonin depletion attenuates ventilation and body temperature in young but not adult Tph2 knockout rats
Abstract
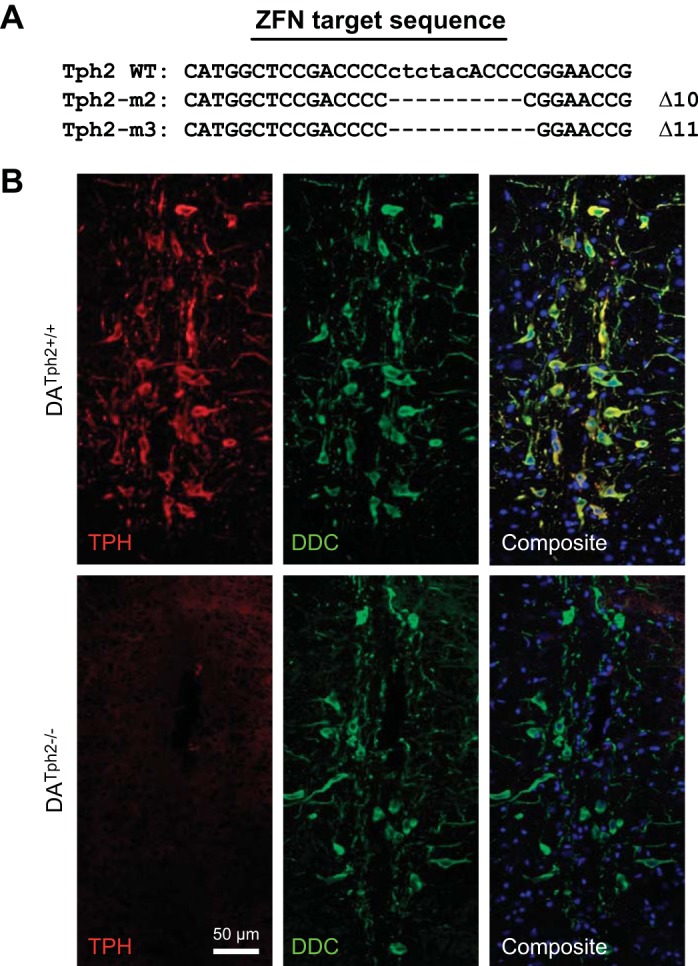
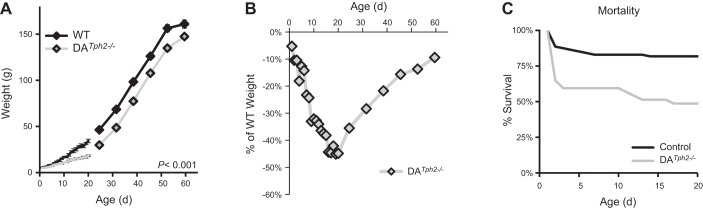
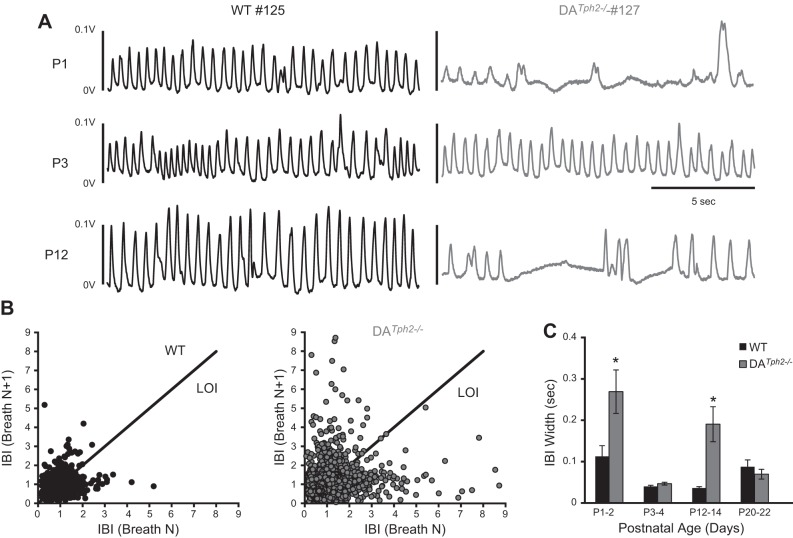
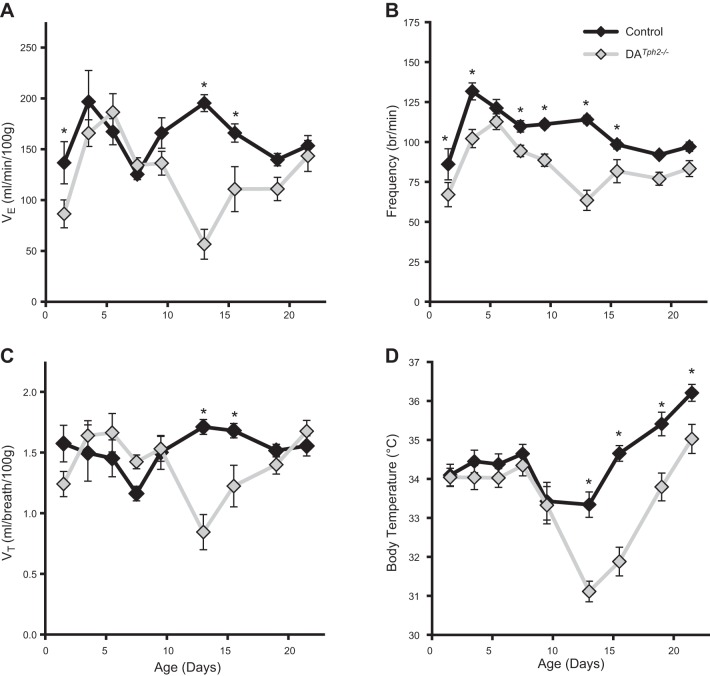
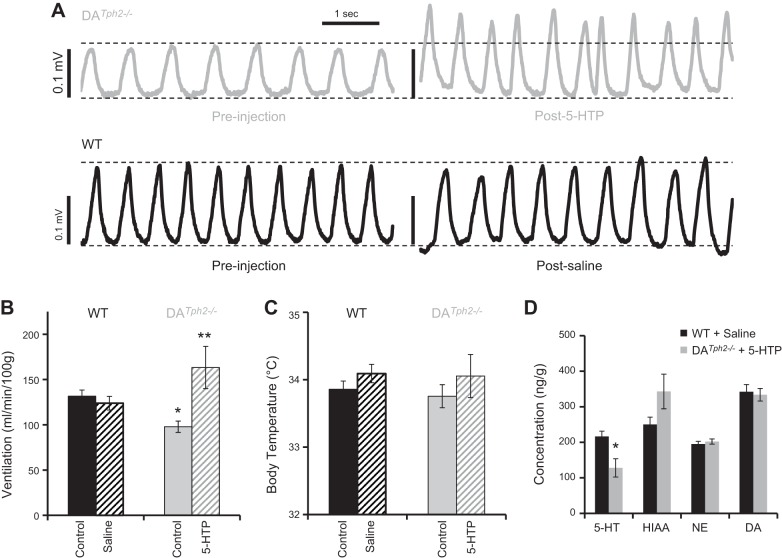
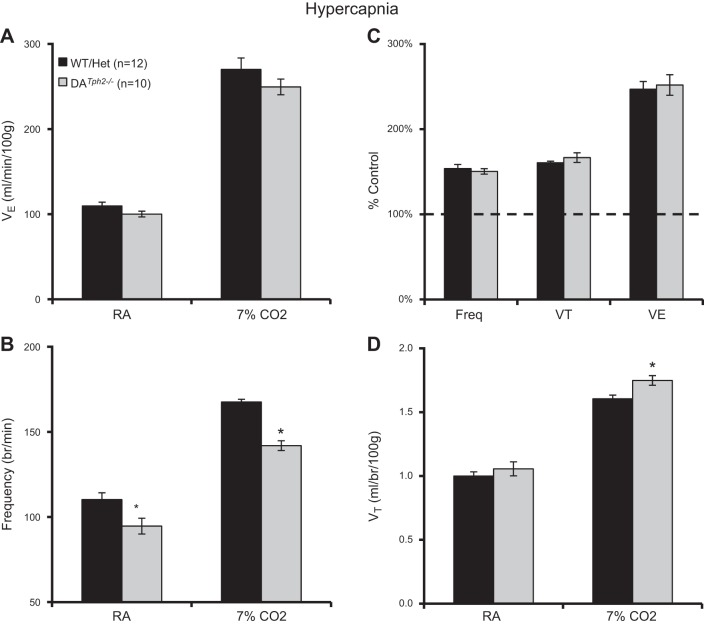
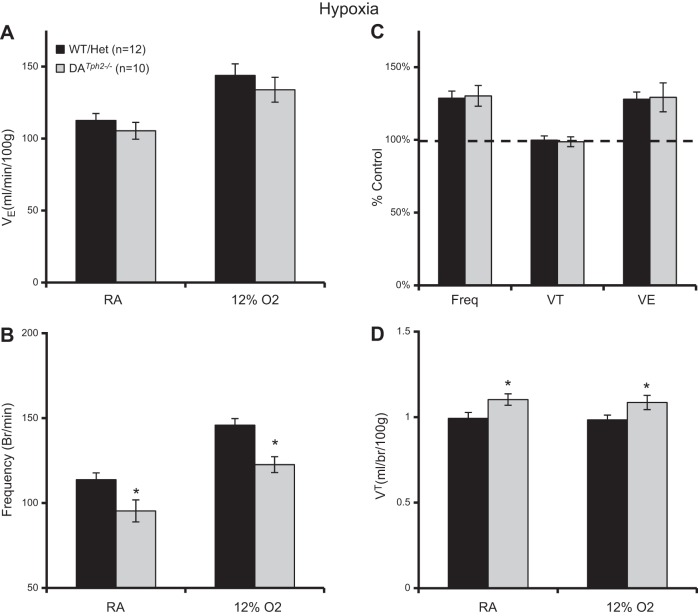
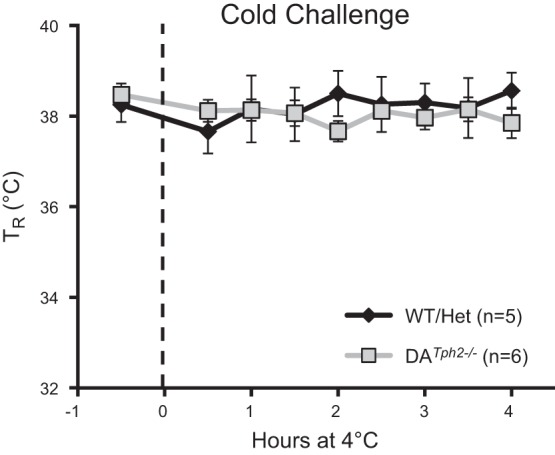
Genetic deletion of brain serotonin (5-HT) neurons in mice leads to ventilatory deficits and increased neonatal mortality during development. However, it is unclear if the loss of the 5-HT neurons or the loss of the neurochemical 5-HT led to the observed physiologic deficits. Herein, we generated a mutant rat model with constitutive central nervous system (CNS) 5-HT depletion by mutation of the tryptophan hydroxylase 2 (Tph2) gene in dark agouti (DA(Tph2-/-)) rats. DA(Tph2-/-) rats lacked TPH immunoreactivity and brain 5-HT but retain dopa decarboxylase-expressing raphe neurons. Mutant rats were also smaller, had relatively high mortality (∼50%), and compared with controls had reduced room air ventilation and body temperatures at specific postnatal ages. In adult rats, breathing at rest and hypoxic and hypercapnic chemoreflexes were unaltered in adult male and female DA(Tph2-/-) rats. Body temperature was also maintained in adult DA(Tph2-/-) rats exposed to 4°C, indicating unaltered ventilatory and/or thermoregulatory control mechanisms. Finally, DA(Tph2-/-) rats treated with the 5-HT precursor 5-hydroxytryptophan (5-HTP) partially restored CNS 5-HT and showed increased ventilation (P < 0.05) at a developmental age when it was otherwise attenuated in the mutants. We conclude that constitutive CNS production of 5-HT is critically important to fundamental homeostatic control systems for breathing and temperature during postnatal development in the rat.
Keywords: apnea; chemoreflex; control of breathing; neonate; serotonin.
Copyright © 2016 the American Physiological Society.
Figures
Similar articles
-
Active sleep unmasks apnea and delayed arousal in infant rat pups lacking central serotonin.J Appl Physiol (1985). 2017 Oct 1;123(4):825-834. doi: 10.1152/japplphysiol.00439.2017. Epub 2017 Aug 3. J Appl Physiol (1985). 2017. PMID: 28775068 Free PMC article.
-
Effects of Serotonin and Slow-Release 5-Hydroxytryptophan on Gastrointestinal Motility in a Mouse Model of Depression.Gastroenterology. 2019 Aug;157(2):507-521.e4. doi: 10.1053/j.gastro.2019.04.022. Epub 2019 May 7. Gastroenterology. 2019. PMID: 31071306 Free PMC article.
-
Impacts of brain serotonin deficiency following Tph2 inactivation on development and raphe neuron serotonergic specification.PLoS One. 2012;7(8):e43157. doi: 10.1371/journal.pone.0043157. Epub 2012 Aug 17. PLoS One. 2012. PMID: 22912815 Free PMC article.
-
Serotonergic neurotransmission manipulation for the understanding of brain development and function: Learning from Tph2 genetic models.Biochimie. 2019 Jun;161:3-14. doi: 10.1016/j.biochi.2018.11.016. Epub 2018 Dec 1. Biochimie. 2019. PMID: 30513372 Review.
-
Life without brain serotonin: reevaluation of serotonin function with mice deficient in brain serotonin synthesis.Behav Brain Res. 2015 Jan 15;277:78-88. doi: 10.1016/j.bbr.2014.06.005. Epub 2014 Jun 11. Behav Brain Res. 2015. PMID: 24928769 Review.
Cited by
-
Central serotonin and the control of arterial blood pressure and heart rate in infant rats: influence of sleep state and sex.Am J Physiol Regul Integr Comp Physiol. 2018 Feb 1;314(2):R313-R321. doi: 10.1152/ajpregu.00321.2017. Epub 2017 Nov 14. Am J Physiol Regul Integr Comp Physiol. 2018. PMID: 29046318 Free PMC article.
-
Constitutive depletion of brain serotonin differentially affects rats' social and cognitive abilities.iScience. 2023 Jan 18;26(2):105998. doi: 10.1016/j.isci.2023.105998. eCollection 2023 Feb 17. iScience. 2023. PMID: 36798444 Free PMC article.
-
TPH2 Deficiency Influences Neuroplastic Mechanisms and Alters the Response to an Acute Stress in a Sex Specific Manner.Front Mol Neurosci. 2018 Oct 30;11:389. doi: 10.3389/fnmol.2018.00389. eCollection 2018. Front Mol Neurosci. 2018. PMID: 30425618 Free PMC article.
-
Tryptophan hydroxylase 2 deficiency alters autism-related behavioural phenotypes in rats.Sci Rep. 2025 Jul 1;15(1):20522. doi: 10.1038/s41598-025-05684-9. Sci Rep. 2025. PMID: 40594285 Free PMC article.
-
Mortality and ventilatory effects of central serotonin deficiency during postnatal development depend on age but not sex.Physiol Rep. 2021 Jul;9(13):e14946. doi: 10.14814/phy2.14946. Physiol Rep. 2021. PMID: 34228894 Free PMC article.
References
-
- Alenina N, Klempin F. The role of serotonin in adult hippocampal neurogenesis. Behav Brain Res 277: 49–57, 2015. - PubMed
-
- Cummings K, Commons K, Nattie EE. Destruction of serotonergic neurons in the neonatal rat brainstem decreases breathing and exacerbates apnea-induced bradycardia; evidence for a role for 5-HT signaling in Sudden Infant Death Syndrome (SIDS). Program No. 383.12. Neuroscience 2008 Abstracts. Washington, DC: Society for Neuroscience, 2008, online.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources