

Lipid Droplet Formation, Their Localization and Dynamics during Leishmania major Macrophage Infection
- PMID: 26871576
- PMCID: PMC4752496
- DOI: 10.1371/journal.pone.0148640
Lipid Droplet Formation, Their Localization and Dynamics during Leishmania major Macrophage Infection
Abstract
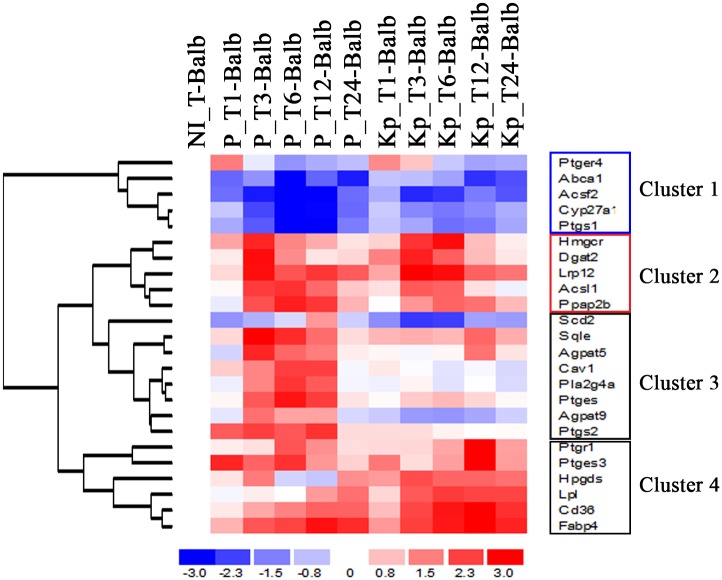
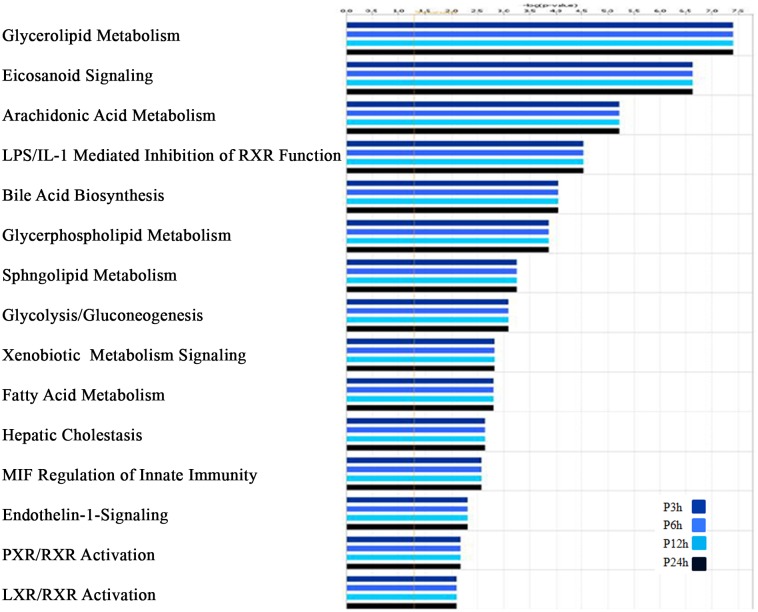
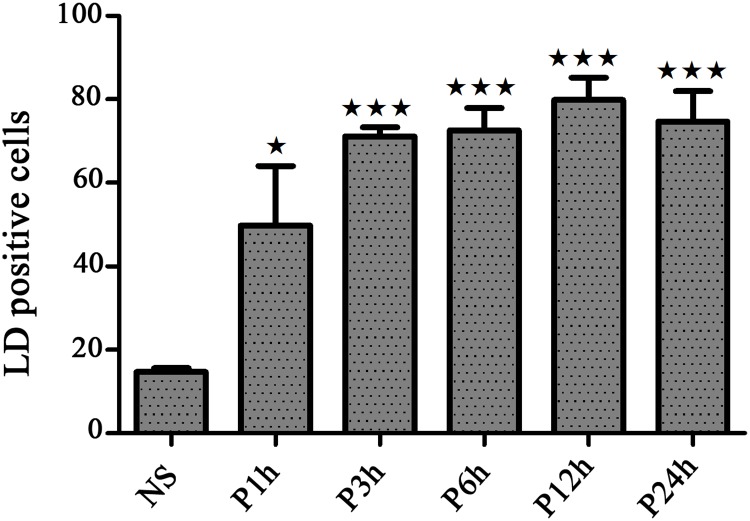
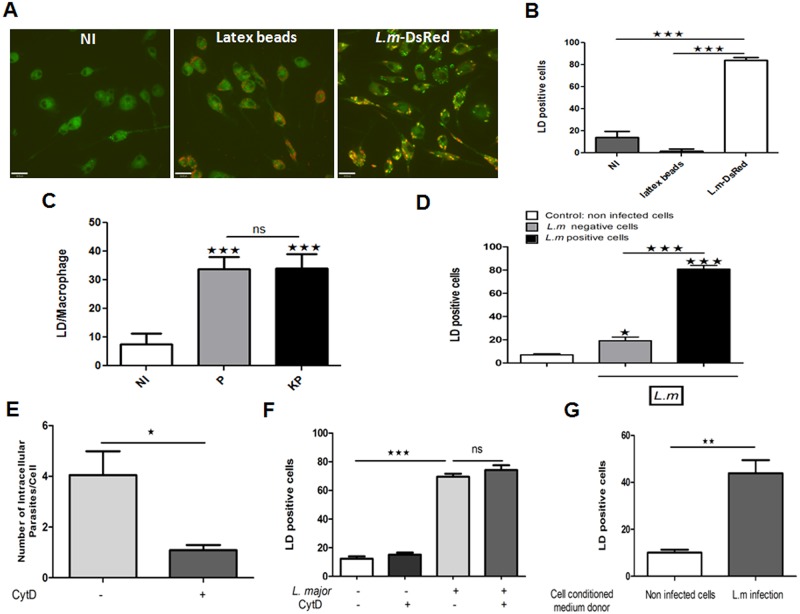
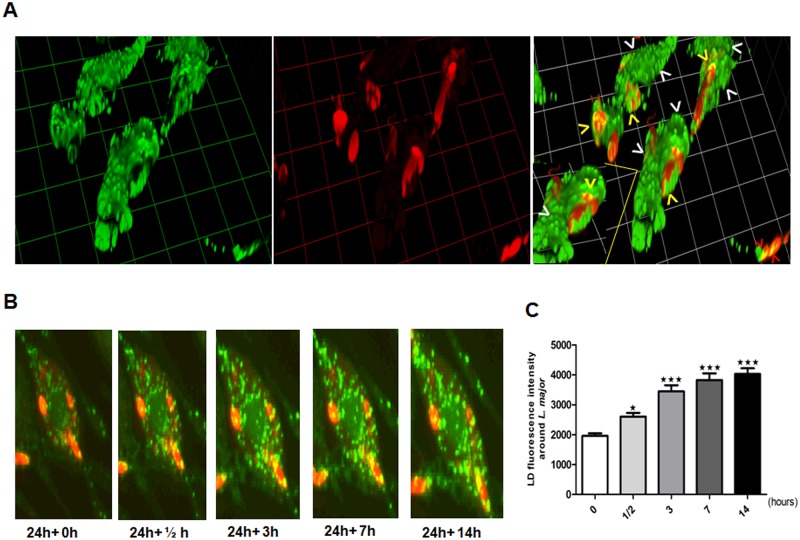
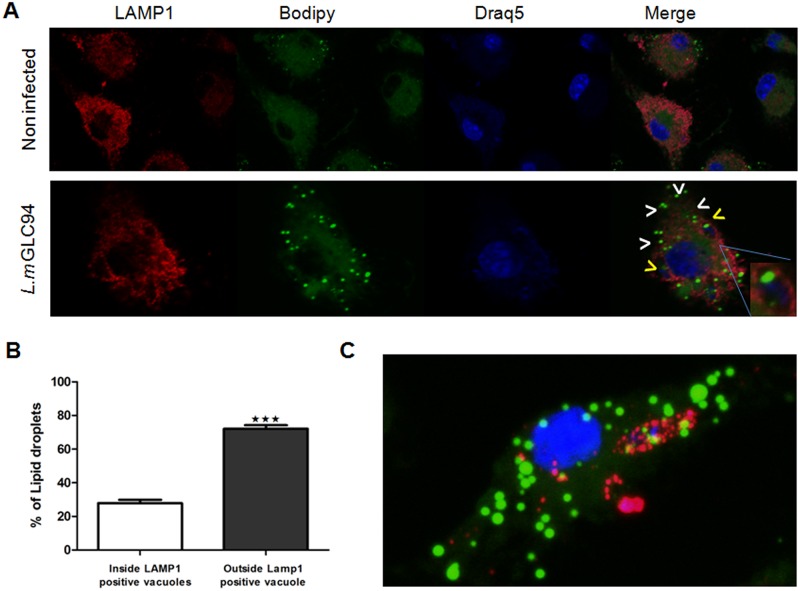
Leishmania, the causative agent of vector-borne diseases, known as leishmaniases, is an obligate intracellular parasite within mammalian hosts. The outcome of infection depends largely on the activation status of macrophages, the first line of mammalian defense and the major target cells for parasite replication. Understanding the strategies developed by the parasite to circumvent macrophage defense mechanisms and to survive within those cells help defining novel therapeutic approaches for leishmaniasis. We previously showed the formation of lipid droplets (LDs) in L. major infected macrophages. Here, we provide novel insights on the origin of the formed LDs by determining their cellular distribution and to what extent these high-energy sources are directed to the proximity of Leishmania parasites. We show that the ability of L. major to trigger macrophage LD accumulation is independent of parasite viability and uptake and can also be observed in non-infected cells through paracrine stimuli suggesting that LD formation is from cellular origin. The accumulation of LDs is demonstrated using confocal microscopy and live-cell imagin in parasite-free cytoplasmic region of the host cell, but also promptly recruited to the proximity of Leishmania parasites. Indeed LDs are observed inside parasitophorous vacuole and in parasite cytoplasm suggesting that Leishmania parasites besides producing their own LDs, may take advantage of these high energy sources. Otherwise, these LDs may help cells defending against parasitic infection. These metabolic changes, rising as common features during the last years, occur in host cells infected by a large number of pathogens and seem to play an important role in pathogenesis. Understanding how Leishmania parasites and different pathogens exploit this LD accumulation will help us define the common mechanism used by these different pathogens to manipulate and/or take advantage of this high-energy source.
Conflict of interest statement
Figures
References
-
- WHO. Leishmaniasis [Internet]. 2015 p. http://www.who.int/mediacentre/factsheets/fs375/en.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
