

The Mechanistic Underpinnings of an ago1-Mediated, Environmentally Dependent, and Stochastic Phenotype
- PMID: 26872948
- PMCID: PMC4825122
- DOI: 10.1104/pp.15.01928
The Mechanistic Underpinnings of an ago1-Mediated, Environmentally Dependent, and Stochastic Phenotype
Abstract
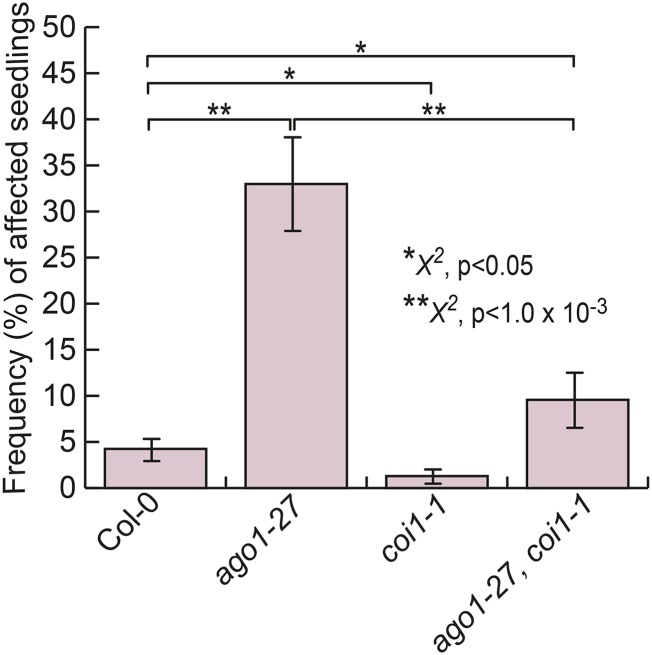
The crucial role of microRNAs in plant development is exceedingly well supported; their importance in environmental robustness is studied in less detail. Here, we describe a novel, environmentally dependent phenotype in hypomorphic argonaute1 (ago1) mutants and uncover its mechanistic underpinnings in Arabidopsis (Arabidopsis thaliana). AGO1 is a key player in microRNA-mediated gene regulation. We observed transparent lesions on embryonic leaves of ago1 mutant seedlings. These lesions increased in frequency in full-spectrum light. Notably, the lesion phenotype was most environmentally responsive in ago1-27 mutants. This allele is thought to primarily affect translational repression, which has been linked with the response to environmental perturbation. Using several lines of evidence, we found that these lesions represent dead and dying tissues due to an aberrant hypersensitive response. Although all three canonical defense hormone pathways (salicylic acid, jasmonate, and jasmonate/ethylene pathways) were up-regulated in ago1 mutants, we demonstrate that jasmonate perception drives the lesion phenotype. Double mutants of ago1 and coronatine insensitive1, the jasmonate receptor, showed greatly decreased frequency of affected seedlings. The chaperone HEAT SHOCK PROTEIN 90 (HSP90), which maintains phenotypic robustness in the face of environmental perturbations, is known to facilitate AGO1 function. HSP90 perturbation has been shown previously to up-regulate jasmonate signaling and to increase plant resistance to herbivory. Although single HSP90 mutants showed subtly elevated levels of lesions, double mutant analysis disagreed with a simple epistatic model for HSP90 and AGO1 interaction; rather, both appeared to act nonadditively in producing lesions. In summary, our study identifies AGO1 as a major, largely HSP90-independent, factor in providing environmental robustness to plants.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures






References
-
- A-H-Mackerness S, Surplus SL, Blake P, John CF, Buchanan-Wollaston V, Jordan BR, Thomas B (1999) Ultraviolet-B-induced stress and changes in gene expression in Arabidopsis thaliana: role of signalling pathways controlled by jasmonic acid, ethylene and reactive oxygen species. Plant Cell Environ 22: 1413–1423
-
- Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR (1999) EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284: 2148–2152 - PubMed
-
- Anderson JP, Badruzsaufari E, Schenk PM, Manners JM, Desmond OJ, Ehlert C, Maclean DJ, Ebert PR, Kazan K (2004) Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 16: 3460–3479 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
