

Methamphetamine addiction: involvement of CREB and neuroinflammatory signaling pathways
- PMID: 26873080
- PMCID: PMC5627363
- DOI: 10.1007/s00213-016-4235-8
Methamphetamine addiction: involvement of CREB and neuroinflammatory signaling pathways
Abstract
Rationale and objectives: Addiction to psychostimulant methamphetamine (METH) remains a major public health problem in the world. Animal models that use METH self-administration incorporate many features of human drug-taking behavior and are very helpful in elucidating mechanisms underlying METH addiction. These models are also helping to decipher the neurobiological substrates of associated neuropsychiatric complications. This review summarizes our work on the influence of METH self-administration on dopamine systems, transcription and immune responses in the brain.
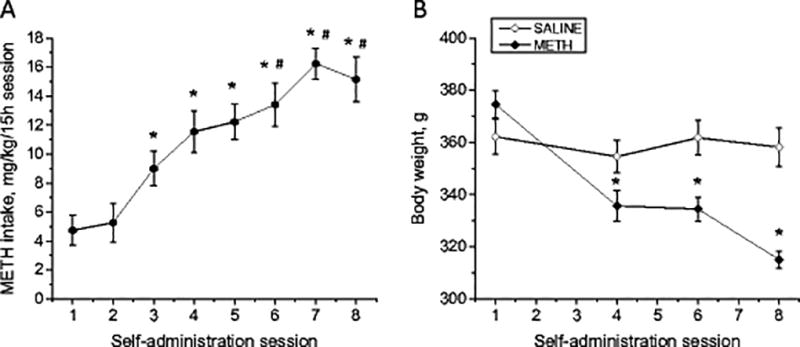
Methods: We used the rat model of METH self-administration with extended access (15 h/day for eight consecutive days) to investigate the effects of voluntary METH intake on the markers of dopamine system integrity and changes in gene expression observed in the brain at 2 h-1 month after cessation of drug exposure.
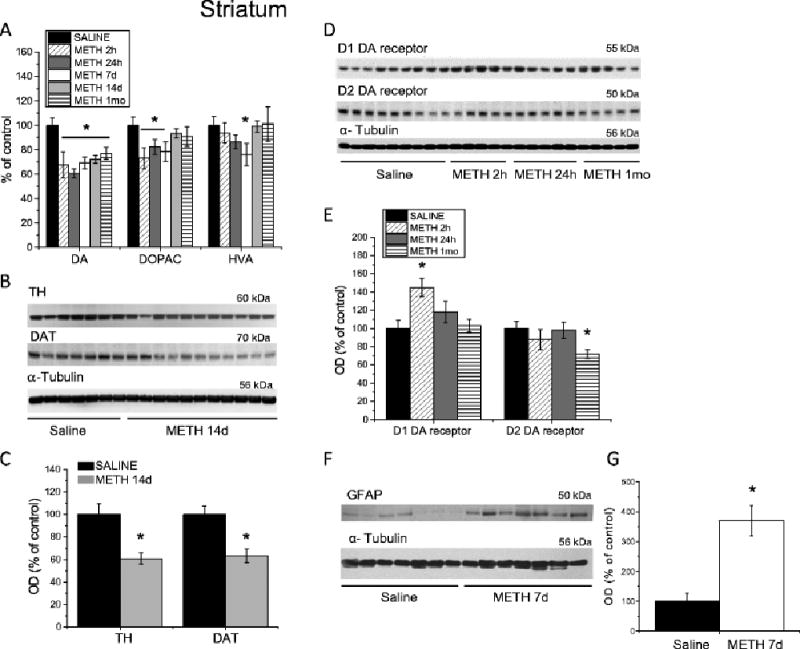
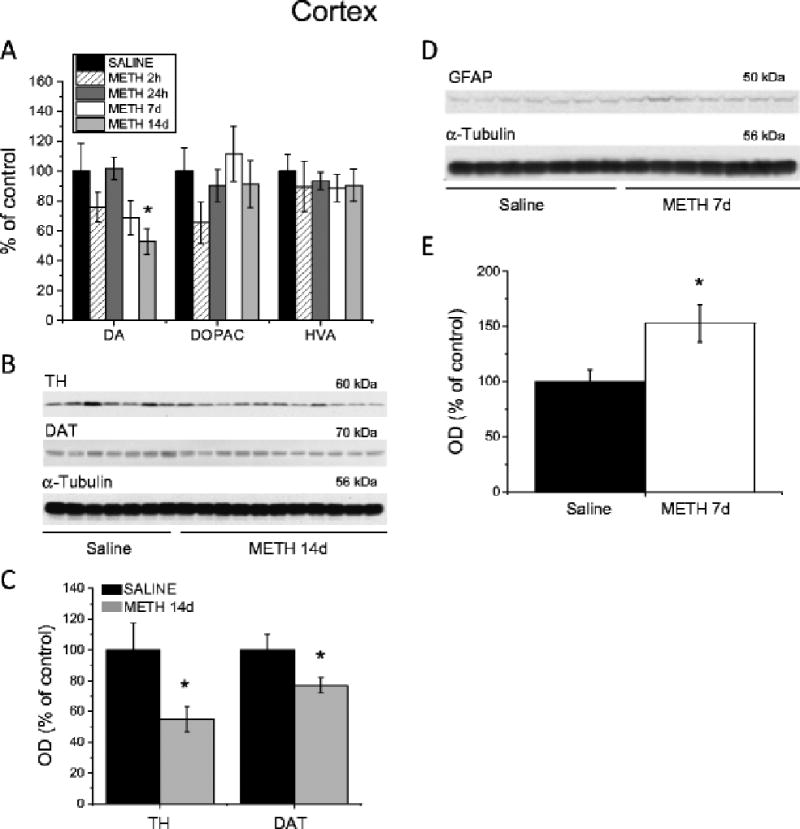
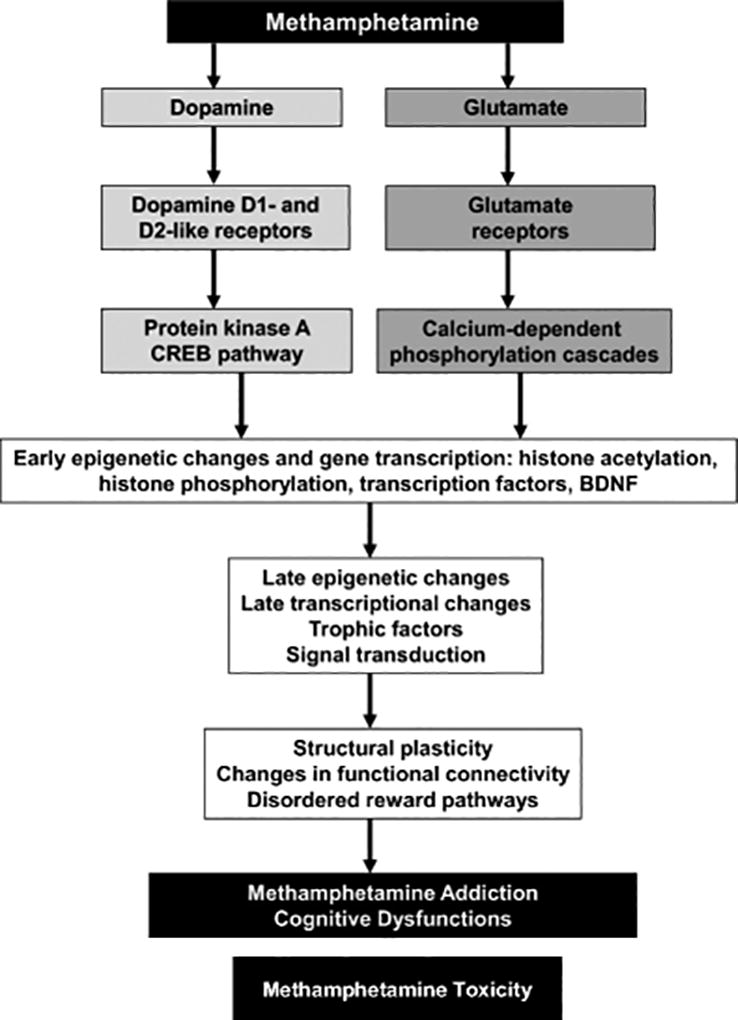
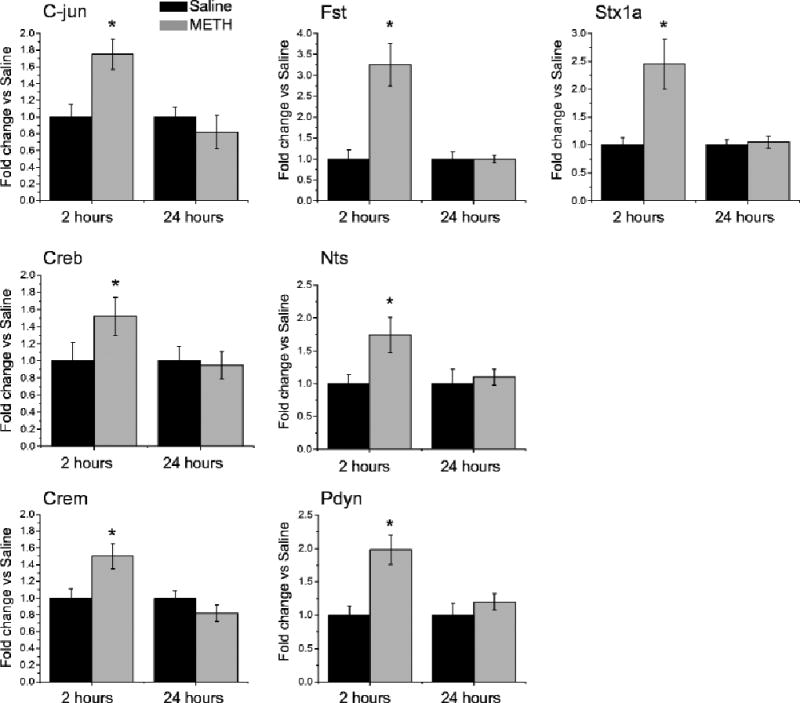
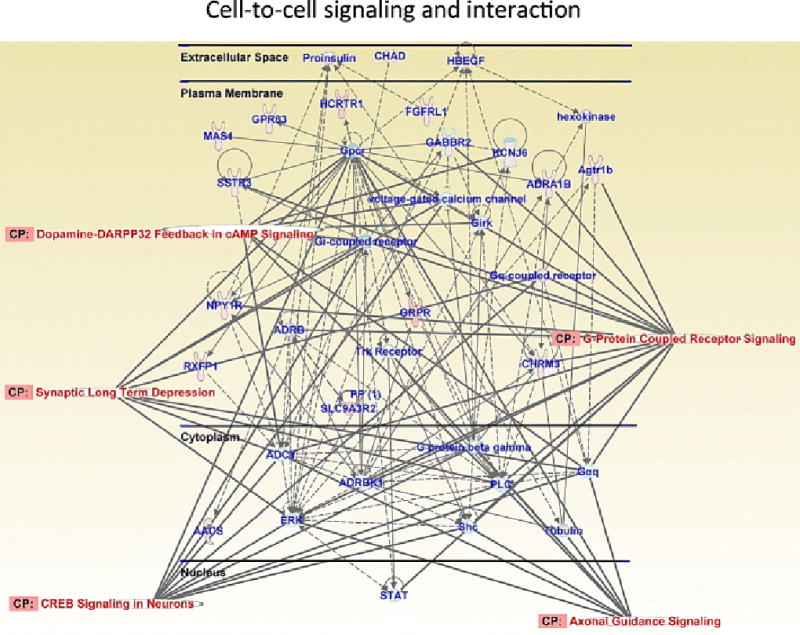
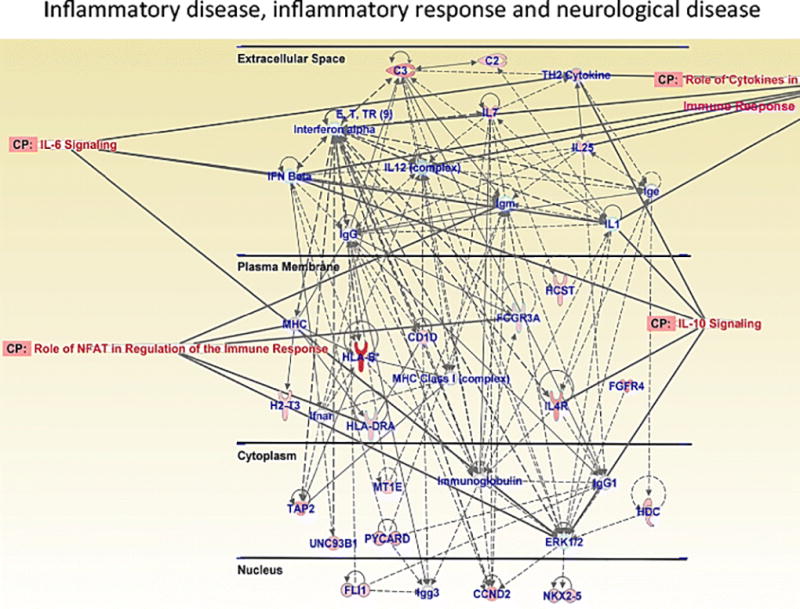
Results: Extended access to METH self-administration caused changes in the rat brain that are consistent with clinical findings reported in neuroimaging and postmortem studies of human METH addicts. In addition, gene expression studies using striatal tissues from METH self-administering rats revealed increased expression of genes involved in cAMP response element binding protein (CREB) signaling pathway and in the activation of neuroinflammatory response in the brain.
Conclusion: These data show an association of METH exposure with activation of neuroplastic and neuroinflammatory cascades in the brain. The neuroplastic changes may be involved in promoting METH addiction. Neuroinflammatory processes in the striatum may underlie cognitive deficits, depression, and parkinsonism reported in METH addicts. Therapeutic approaches that include suppression of neuroinflammation may be beneficial to addicted patients.
Keywords: Cortex; Dopamine; Gene expression; Neuroinflammation; Neurotoxicity; Self-administration; Striatum.
Conflict of interest statement
Figures
Similar articles
-
CREB phosphorylation regulates striatal transcriptional responses in the self-administration model of methamphetamine addiction in the rat.Neurobiol Dis. 2013 Oct;58:132-43. doi: 10.1016/j.nbd.2013.05.009. Epub 2013 May 30. Neurobiol Dis. 2013. PMID: 23726845 Free PMC article.
-
Methamphetamine reduces expression of caveolin-1 in the dorsal striatum: Implication for dysregulation of neuronal function.Neuroscience. 2016 Jul 22;328:147-56. doi: 10.1016/j.neuroscience.2016.04.039. Epub 2016 Apr 30. Neuroscience. 2016. PMID: 27138644 Free PMC article.
-
Methamphetamine induces Shati/Nat8L expression in the mouse nucleus accumbens via CREB- and dopamine D1 receptor-dependent mechanism.PLoS One. 2017 Mar 20;12(3):e0174196. doi: 10.1371/journal.pone.0174196. eCollection 2017. PLoS One. 2017. PMID: 28319198 Free PMC article.
-
Epigenetic landscape of amphetamine and methamphetamine addiction in rodents.Epigenetics. 2015;10(7):574-80. doi: 10.1080/15592294.2015.1055441. Epigenetics. 2015. PMID: 26023847 Free PMC article. Review.
-
Neurotoxicity of methamphetamine: Main effects and mechanisms.Exp Neurol. 2021 Oct;344:113795. doi: 10.1016/j.expneurol.2021.113795. Epub 2021 Jun 26. Exp Neurol. 2021. PMID: 34186102 Free PMC article. Review.
Cited by
-
Selegiline acts as neuroprotective agent against methamphetamine-prompted mood and cognitive related behavior and neurotoxicity in rats: Involvement of CREB/BDNF and Akt/GSK3 signal pathways.Iran J Basic Med Sci. 2020 May;23(5):606-615. doi: 10.22038/ijbms.2020.38827.9221. Iran J Basic Med Sci. 2020. PMID: 32742598 Free PMC article.
-
The Crosstalk Between Neurons and Glia in Methamphetamine-Induced Neuroinflammation.Neurochem Res. 2022 Apr;47(4):872-884. doi: 10.1007/s11064-021-03513-9. Epub 2022 Jan 4. Neurochem Res. 2022. PMID: 34982394 Review.
-
Neuroprotective Effects of Forced Exercise and Bupropion on Chronic Methamphetamine-induced Cognitive Impairment via Modulation of cAMP Response Element-binding Protein/Brain-derived Neurotrophic Factor Signaling Pathway, Oxidative Stress, and Inflammatory Biomarkers in Rats.Adv Biomed Res. 2018 Dec 19;7:151. doi: 10.4103/abr.abr_11_18. eCollection 2018. Adv Biomed Res. 2018. PMID: 30662880 Free PMC article.
-
The Common Denominators of Parkinson's Disease Pathogenesis and Methamphetamine Abuse.Curr Neuropharmacol. 2024;22(13):2113-2156. doi: 10.2174/1570159X21666230907151226. Curr Neuropharmacol. 2024. PMID: 37691228 Free PMC article. Review.
-
mRNA changes in nucleus accumbens related to methamphetamine addiction in mice.Sci Rep. 2016 Nov 21;6:36993. doi: 10.1038/srep36993. Sci Rep. 2016. PMID: 27869204 Free PMC article.
References
-
- Asanuma M, Miyazaki I, Higashi Y, Tsuji T, Ogawa N. Specific gene expression and possible involvement of inflammation in methamphetamine-induced neurotoxicity. Ann NY Acad Sci. 2004;1025:69–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
