

Pyruvate and Metabolic Flexibility: Illuminating a Path Toward Selective Cancer Therapies
- PMID: 26873641
- PMCID: PMC4783264
- DOI: 10.1016/j.tibs.2016.01.002
Pyruvate and Metabolic Flexibility: Illuminating a Path Toward Selective Cancer Therapies
Abstract
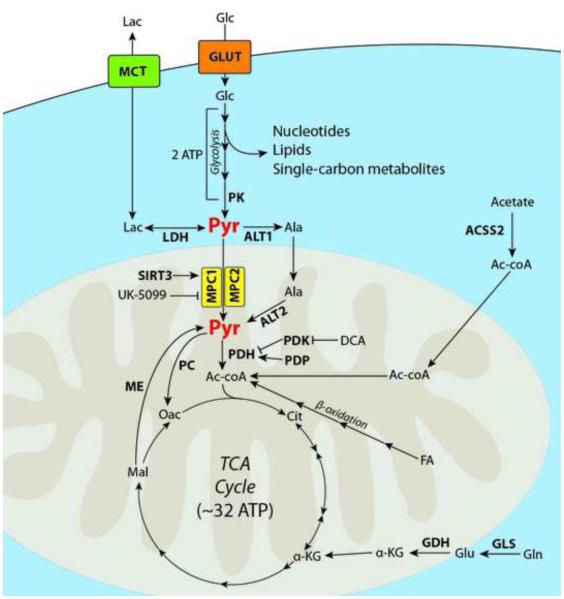
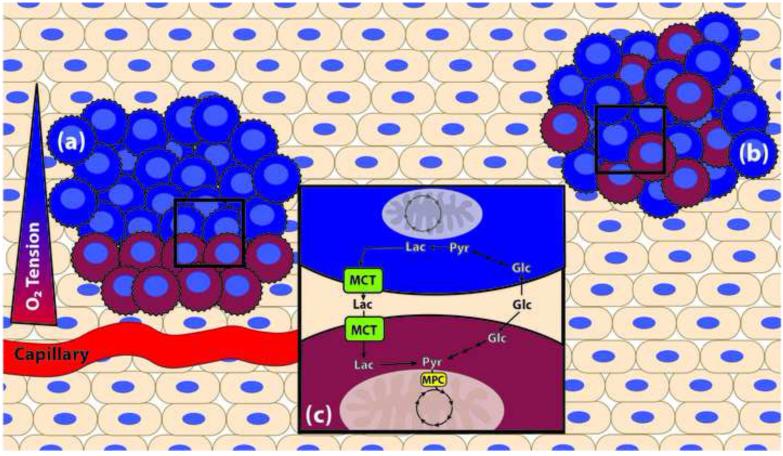
Dysregulated metabolism is an emerging hallmark of cancer, and there is abundant interest in developing therapies to selectively target these aberrant metabolic phenotypes. Sitting at the decision-point between mitochondrial carbohydrate oxidation and aerobic glycolysis (i.e., the 'Warburg effect'), the synthesis and consumption of pyruvate is tightly controlled and is often differentially regulated in cancer cells. This review examines recent efforts toward understanding and targeting mitochondrial pyruvate metabolism, and addresses some of the successes, pitfalls, and significant challenges of metabolic therapy to date.
Keywords: cancer; metabolic flexibility; metabolic heterogeneity; metabolism; pyruvate; stem cells.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Azzi A. memory of Donald Nicholson. IUBMB Life. 2012;648:659–660.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
