

Initiation on the divergent Type I cadicivirus IRES: factor requirements and interactions with the translation apparatus
- PMID: 26873921
- PMCID: PMC4838371
- DOI: 10.1093/nar/gkw074
Initiation on the divergent Type I cadicivirus IRES: factor requirements and interactions with the translation apparatus
Abstract
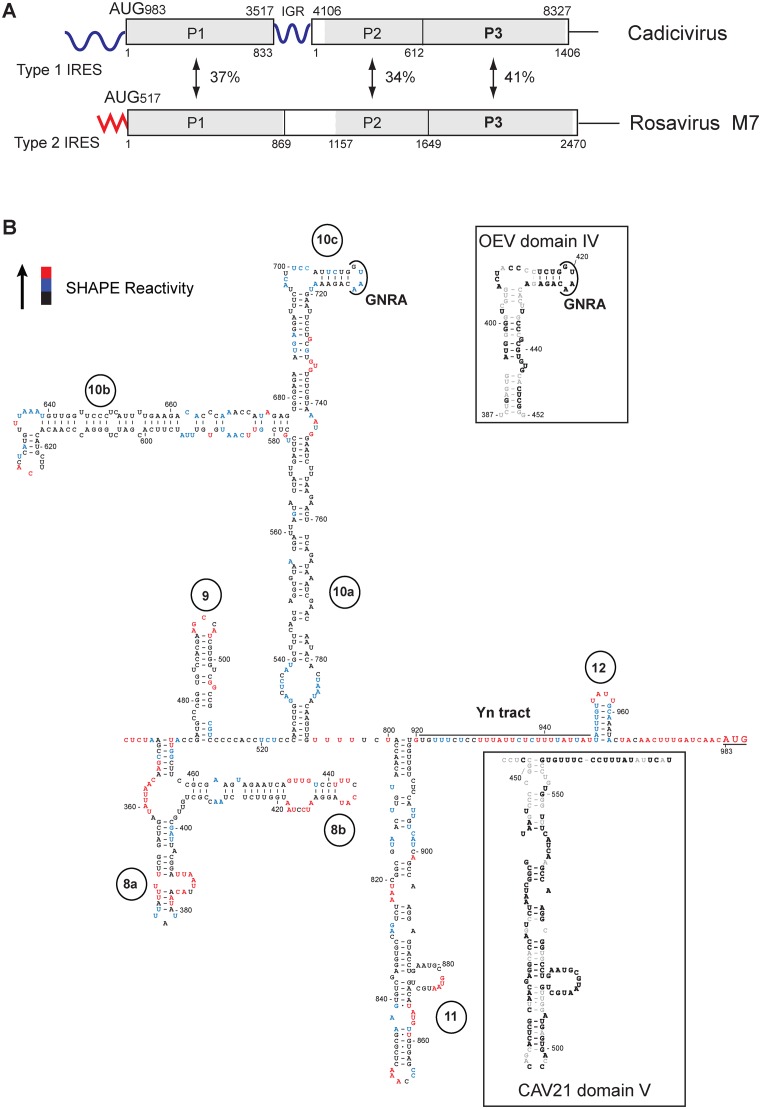
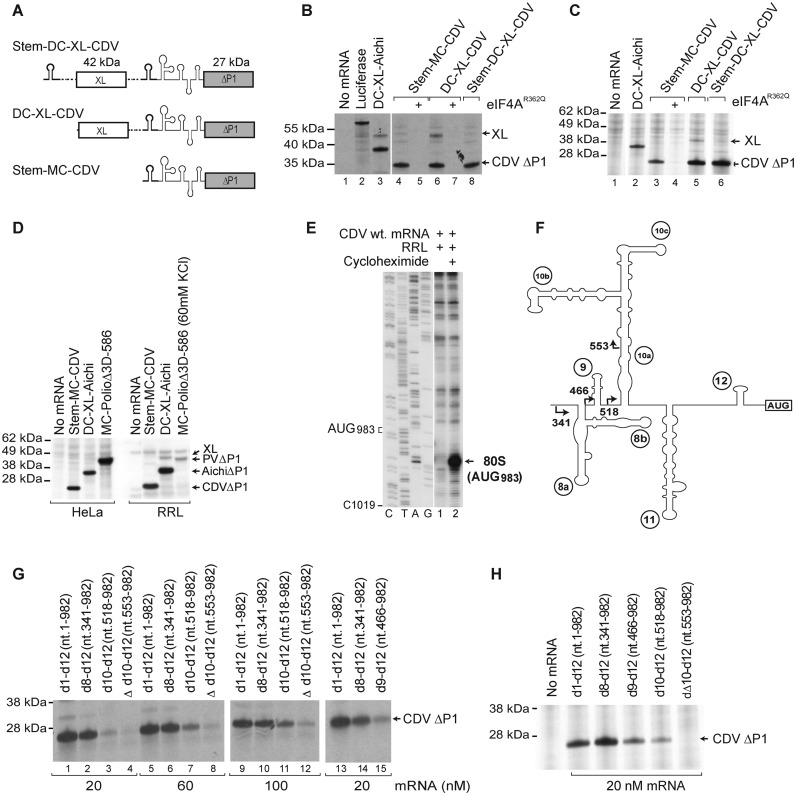
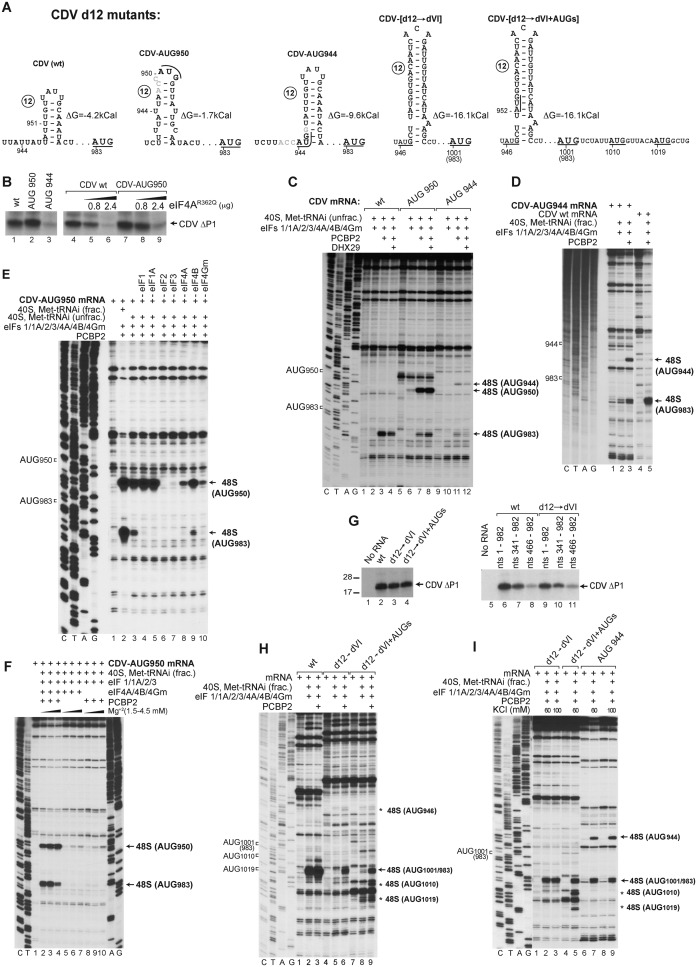
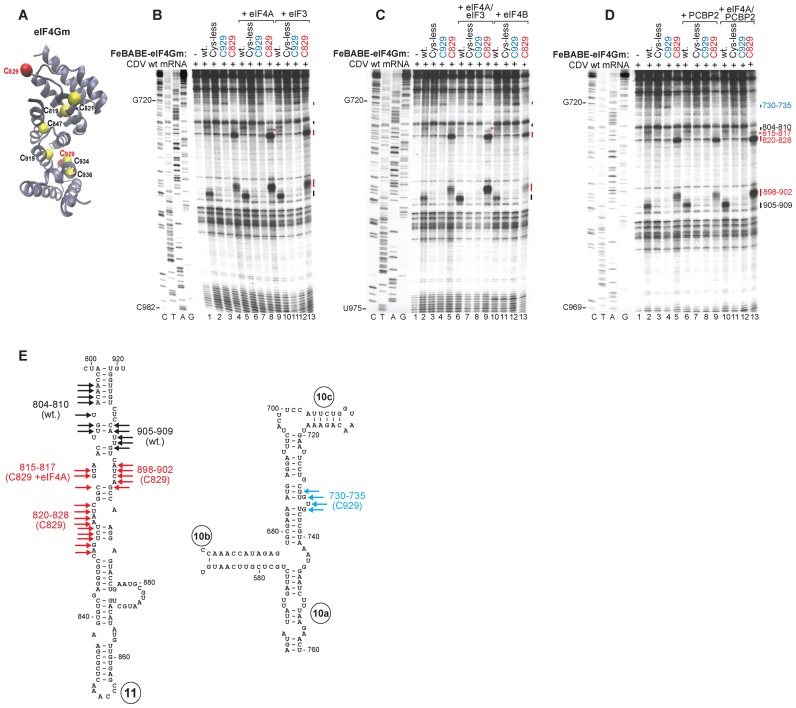
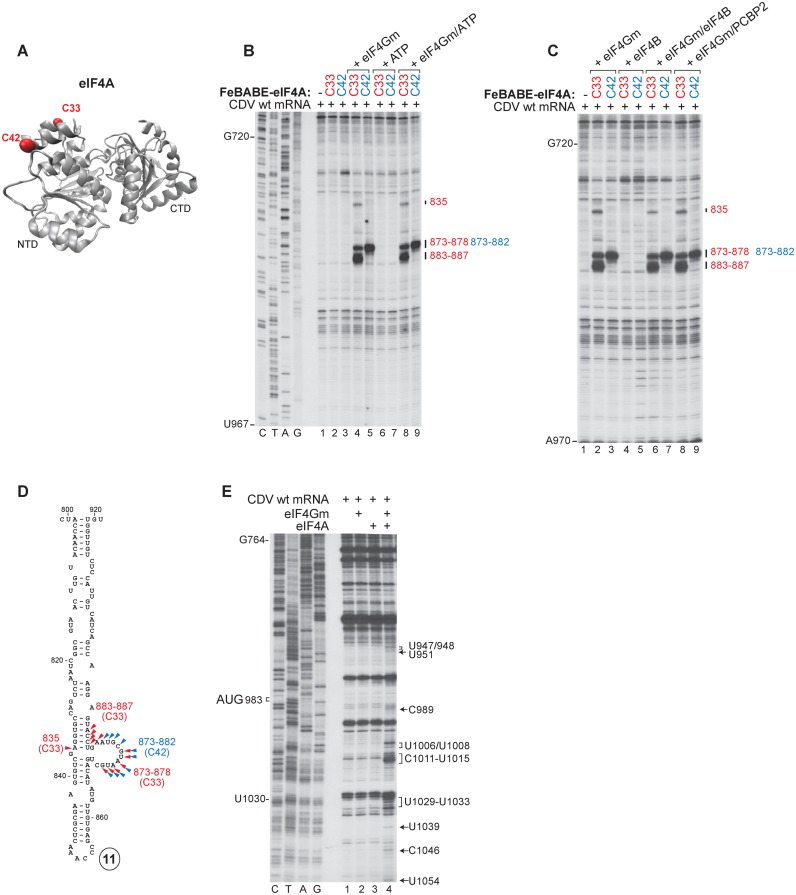
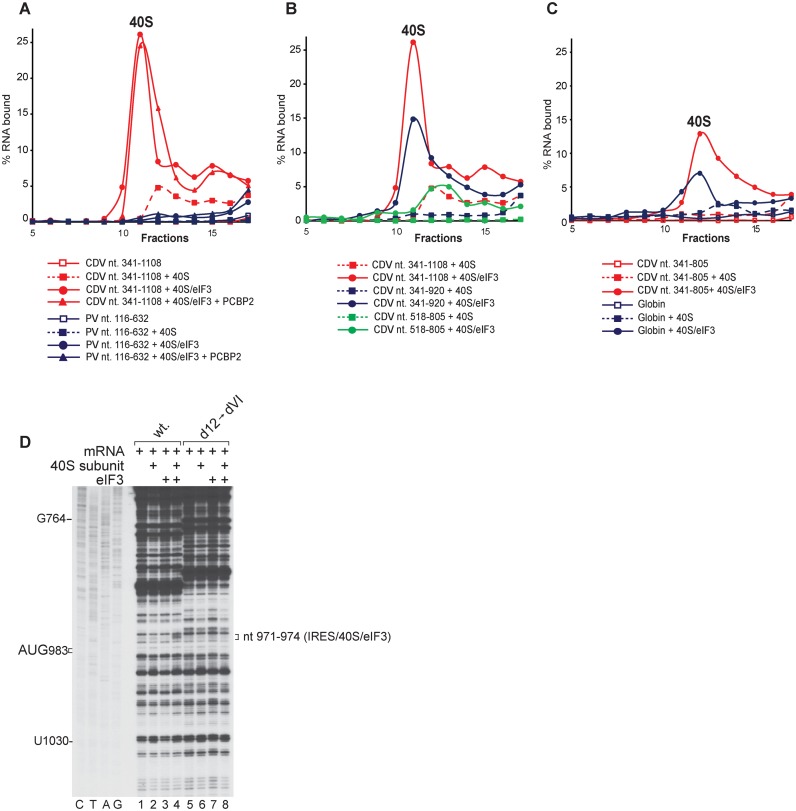
Cadicivirus (CDV) is unique amongst picornaviruses in having a dicistronic genome with internal ribosomal entry sites (IRESs) preceding both open reading frames. Here, we investigated initiation on the 5'-terminal IRES. We report that the 982-nt long 5'UTR comprises 12 domains (d1-d12), five of which (d8-d12, nts 341-950) constitute a divergent Type I IRES. It comprises central elements (the apex of d10, d11 and the following polypyrimidine tract) that are homologous to corresponding elements in canonical Type 1 IRESs, and non-canonical flanking domains (d8, d9 and d12). In vitro reconstitution revealed that as with canonical Type I IRESs, 48S complex formation requires eukaryotic initiation factors (eIFs) 1, 1A, 2, 3, 4A, 4B and 4G, and the poly(C) binding protein 2 (PCBP2), and starts with specific binding of eIF4G/eIF4A to d11. However, in contrast to canonical Type I IRESs, subsequent recruitment of 43S ribosomal complexes does not require direct interaction of their eIF3 constituent with the IRES-bound eIF4G. On the other hand, the CDV IRES forms a 40S/eIF3/IRES ternary complex, with multiple points of contact. These additional interactions with translational components could potentially stimulate recruitment of the 43S complex and alleviate the necessity for direct eIF4G/eIF3 interaction.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures

Similar articles
-
PCBP2 enables the cadicivirus IRES to exploit the function of a conserved GRNA tetraloop to enhance ribosomal initiation complex formation.Nucleic Acids Res. 2016 Nov 16;44(20):9902-9917. doi: 10.1093/nar/gkw609. Epub 2016 Jul 7. Nucleic Acids Res. 2016. PMID: 27387282 Free PMC article.
-
Common conformational changes induced in type 2 picornavirus IRESs by cognate trans-acting factors.Nucleic Acids Res. 2011 Jun;39(11):4851-65. doi: 10.1093/nar/gkr045. Epub 2011 Feb 8. Nucleic Acids Res. 2011. PMID: 21306989 Free PMC article.
-
Hepatitis-C-virus-like internal ribosome entry sites displace eIF3 to gain access to the 40S subunit.Nature. 2013 Nov 28;503(7477):539-43. doi: 10.1038/nature12658. Epub 2013 Nov 3. Nature. 2013. PMID: 24185006 Free PMC article.
-
Hepatitis C Virus Translation Regulation.Int J Mol Sci. 2020 Mar 27;21(7):2328. doi: 10.3390/ijms21072328. Int J Mol Sci. 2020. PMID: 32230899 Free PMC article. Review.
-
Molecular mechanisms of translation initiation in eukaryotes.Proc Natl Acad Sci U S A. 2001 Jun 19;98(13):7029-36. doi: 10.1073/pnas.111145798. Proc Natl Acad Sci U S A. 2001. PMID: 11416183 Free PMC article. Review.
Cited by
-
Horizontal gene transfer as a mechanism for the promiscuous acquisition of distinct classes of IRES by avian caliciviruses.Nucleic Acids Res. 2022 Jan 25;50(2):1052-1068. doi: 10.1093/nar/gkab1243. Nucleic Acids Res. 2022. PMID: 34928389 Free PMC article.
-
eIF4G2 balances its own mRNA translation via a PCBP2-based feedback loop.RNA. 2019 Jul;25(7):757-767. doi: 10.1261/rna.065623.118. Epub 2019 Apr 22. RNA. 2019. PMID: 31010886 Free PMC article.
-
PCBP2 enables the cadicivirus IRES to exploit the function of a conserved GRNA tetraloop to enhance ribosomal initiation complex formation.Nucleic Acids Res. 2016 Nov 16;44(20):9902-9917. doi: 10.1093/nar/gkw609. Epub 2016 Jul 7. Nucleic Acids Res. 2016. PMID: 27387282 Free PMC article.
-
Deconstructing internal ribosome entry site elements: an update of structural motifs and functional divergences.Open Biol. 2018 Nov 28;8(11):180155. doi: 10.1098/rsob.180155. Open Biol. 2018. PMID: 30487301 Free PMC article. Review.
-
Advances and Breakthroughs in IRES-Directed Translation and Replication of Picornaviruses.mBio. 2023 Apr 25;14(2):e0035823. doi: 10.1128/mbio.00358-23. Epub 2023 Mar 20. mBio. 2023. PMID: 36939331 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
