

Synthetic biology to access and expand nature's chemical diversity
- PMID: 26876034
- PMCID: PMC5048682
- DOI: 10.1038/nrmicro.2015.24
Synthetic biology to access and expand nature's chemical diversity
Abstract
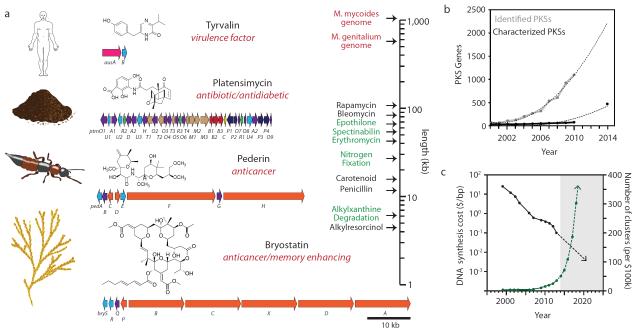
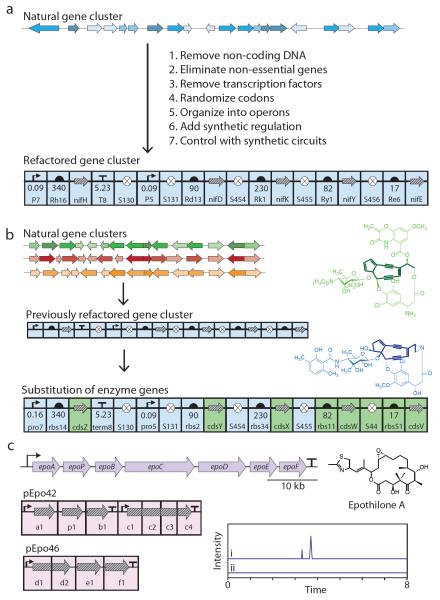
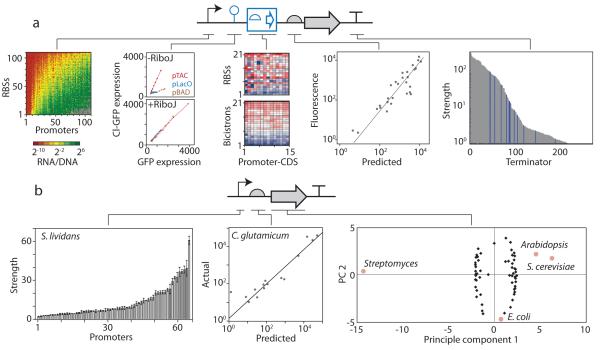
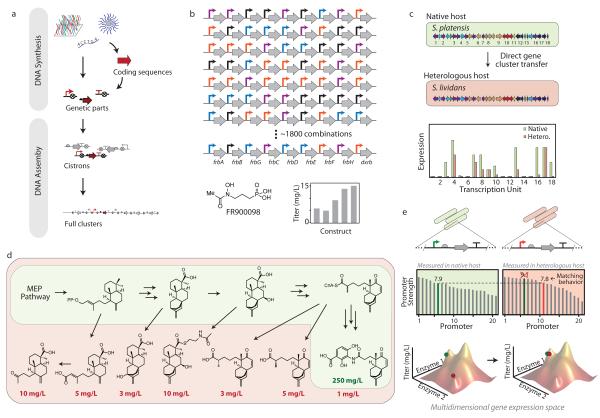
Bacterial genomes encode the biosynthetic potential to produce hundreds of thousands of complex molecules with diverse applications, from medicine to agriculture and materials. Accessing these natural products promises to reinvigorate drug discovery pipelines and provide novel routes to synthesize complex chemicals. The pathways leading to the production of these molecules often comprise dozens of genes spanning large areas of the genome and are controlled by complex regulatory networks with some of the most interesting molecules being produced by non-model organisms. In this Review, we discuss how advances in synthetic biology--including novel DNA construction technologies, the use of genetic parts for the precise control of expression and for synthetic regulatory circuits--and multiplexed genome engineering can be used to optimize the design and synthesis of pathways that produce natural products.
Figures
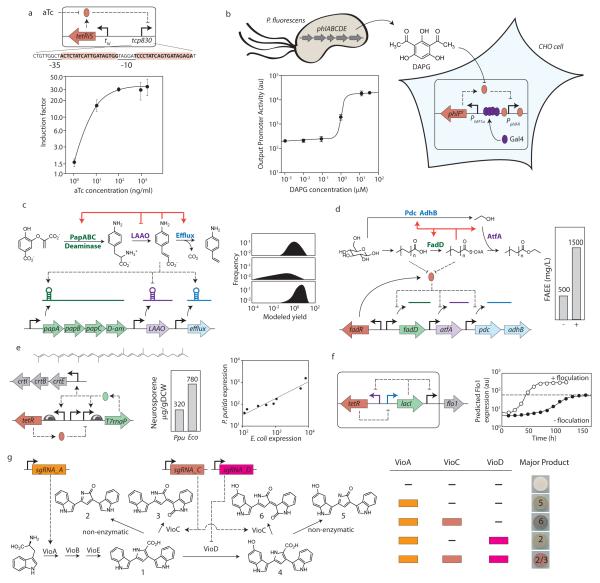
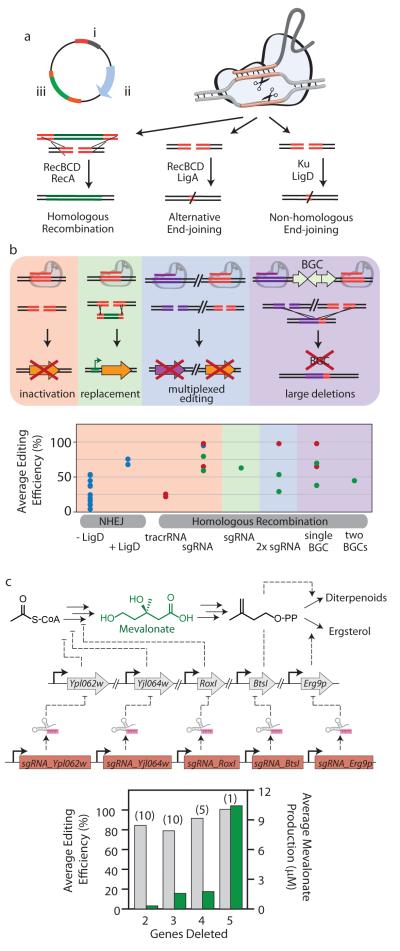
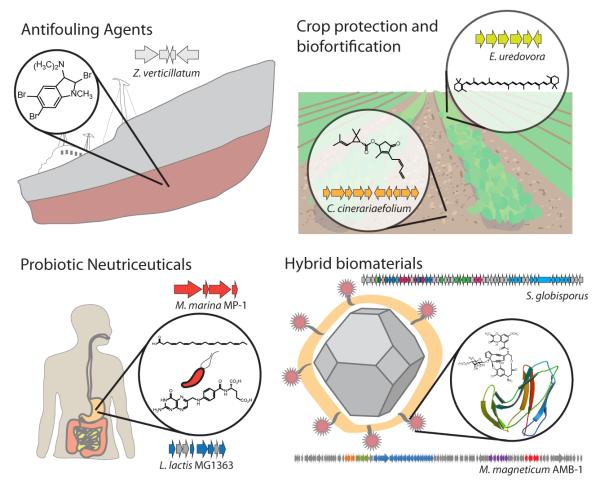
References
-
- Clardy J, Fischbach MA, Walsh CT. New antibiotics from bacterial natural products. Nat. Biotechnol. 2006;24:1541–1550. - PubMed
-
- Cragg GM, Grothaus PG, Newman DJ. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009;109:3012–3043. - PubMed
-
- Mann J. Natural products as immunosuppressive agents. Nat. Prod. Rep. 2001;18:417–430. - PubMed
-
- Demain AL. Importance of microbial natural products and the need to revitalize their discovery. J. Ind. Microbiol. Biotechnol. 2014;41:185–201. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
