

Cyclin-dependent kinase 2 protects podocytes from apoptosis
- PMID: 26876672
- PMCID: PMC4753499
- DOI: 10.1038/srep21664
Cyclin-dependent kinase 2 protects podocytes from apoptosis
Abstract
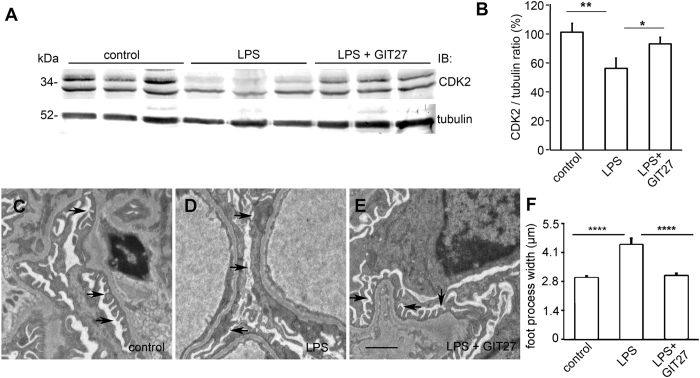
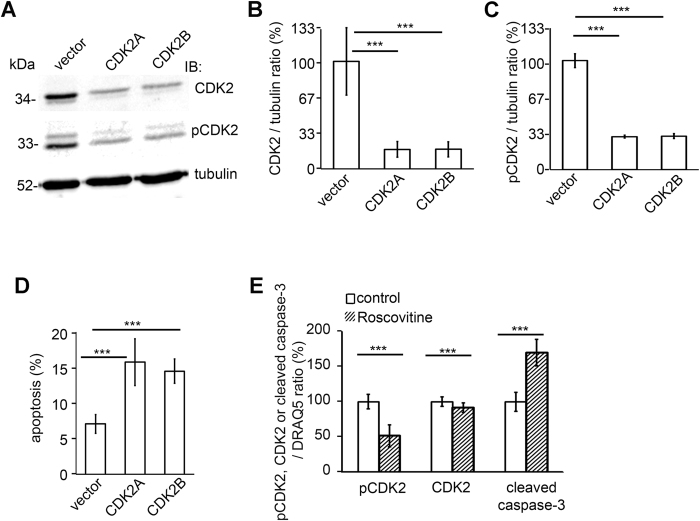
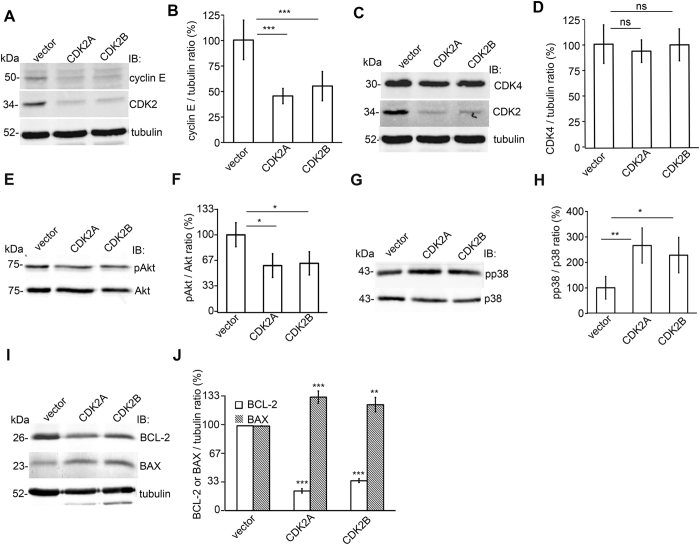
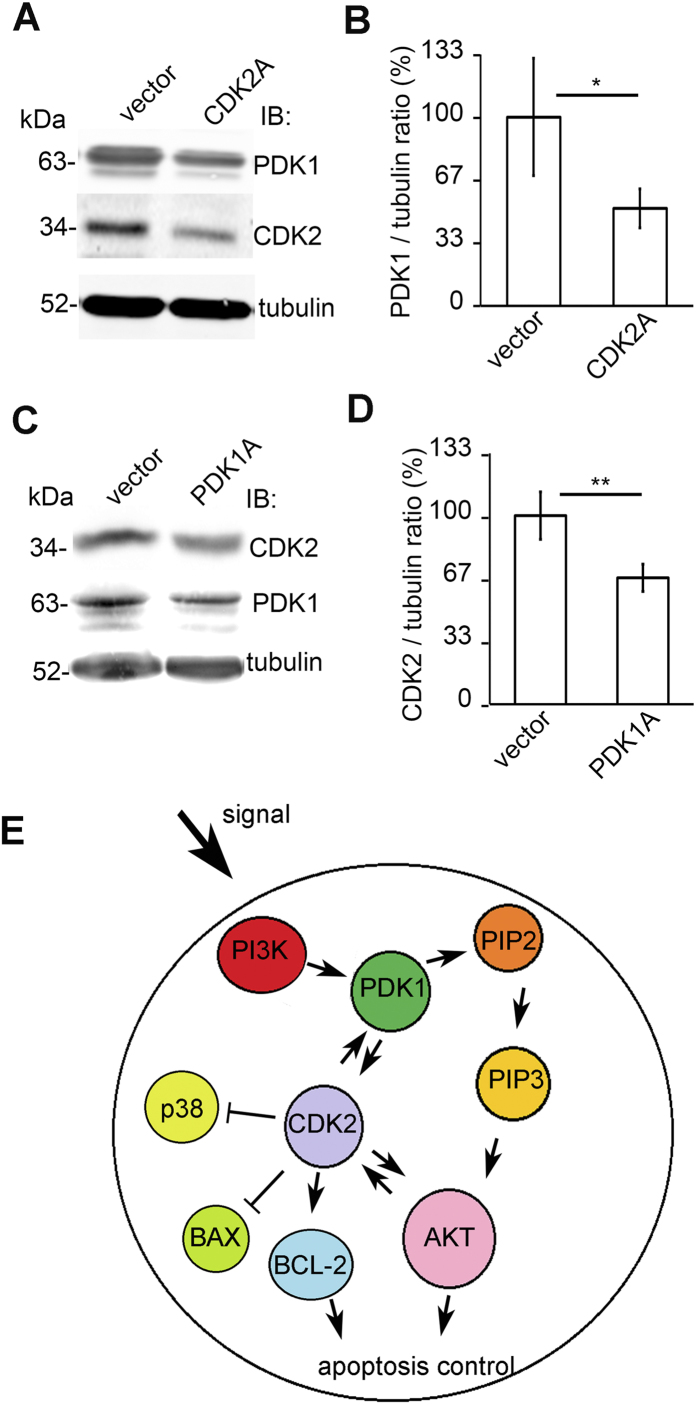
Loss of podocytes is an early feature of diabetic nephropathy (DN) and predicts its progression. We found that treatment of podocytes with sera from normoalbuminuric type 1 diabetes patients with high lipopolysaccharide (LPS) activity, known to predict progression of DN, downregulated CDK2 (cyclin-dependent kinase 2). LPS-treatment of mice also reduced CDK2 expression. LPS-induced downregulation of CDK2 was prevented in vitro and in vivo by inhibiting the Toll-like receptor (TLR) pathway using immunomodulatory agent GIT27. We also observed that CDK2 is downregulated in the glomeruli of obese Zucker rats before the onset of proteinuria. Knockdown of CDK2, or inhibiting its activity with roscovitine in podocytes increased apoptosis. CDK2 knockdown also reduced expression of PDK1, an activator of the cell survival kinase Akt, and reduced Akt phosphorylation. This suggests that CDK2 regulates the activity of the cell survival pathway via PDK1. Furthermore, PDK1 knockdown reduced the expression of CDK2 suggesting a regulatory loop between CDK2 and PDK1. Collectively, our data show that CDK2 protects podocytes from apoptosis and that reduced expression of CDK2 associates with the development of DN. Preventing downregulation of CDK2 by blocking the TLR pathway with GIT27 may provide a means to prevent podocyte apoptosis and progression of DN.
Conflict of interest statement
P-H.G. has received lecture honoraria from Abbot, Boehringer Ingelheim, Cebix, Eli Lilly, Genzyme, Novartis, Novo Nordisk and MSD, and research grants from Eli Lilly and Roche. P-H.G. is an advisory board member for Boehringer Ingelheim, Novartis and Medscape. The authors have no additional financial interests.
Figures
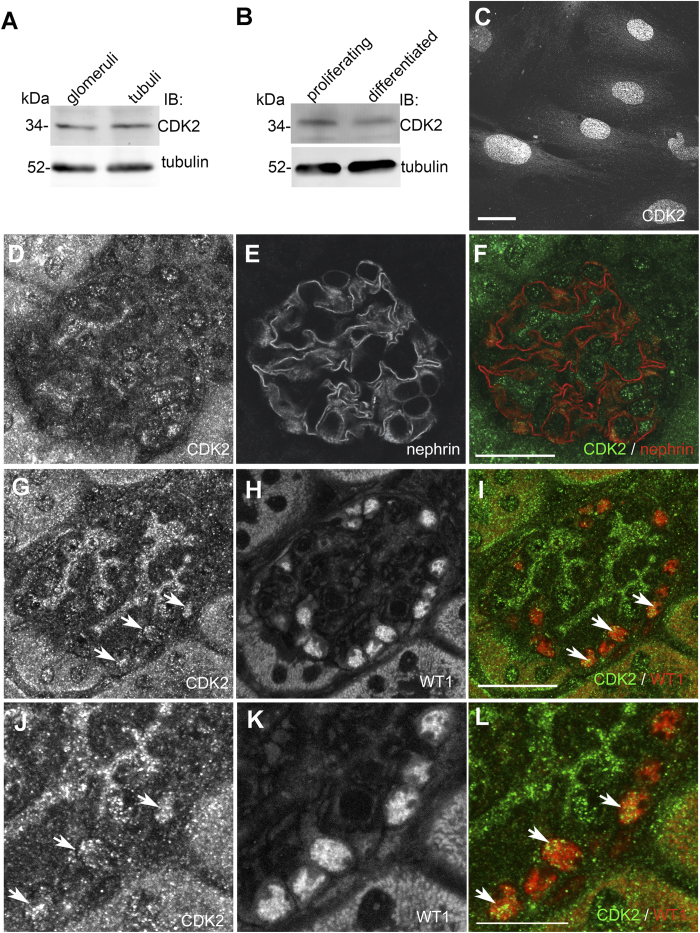
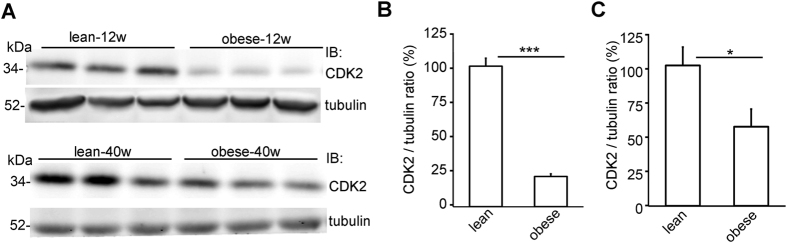
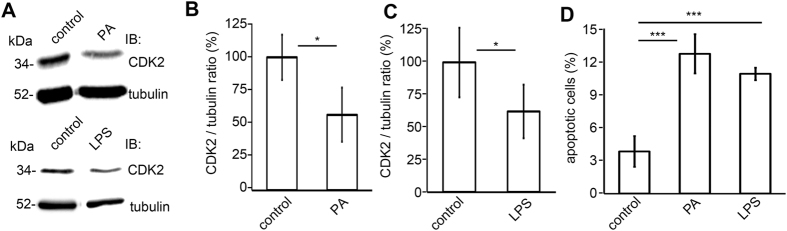
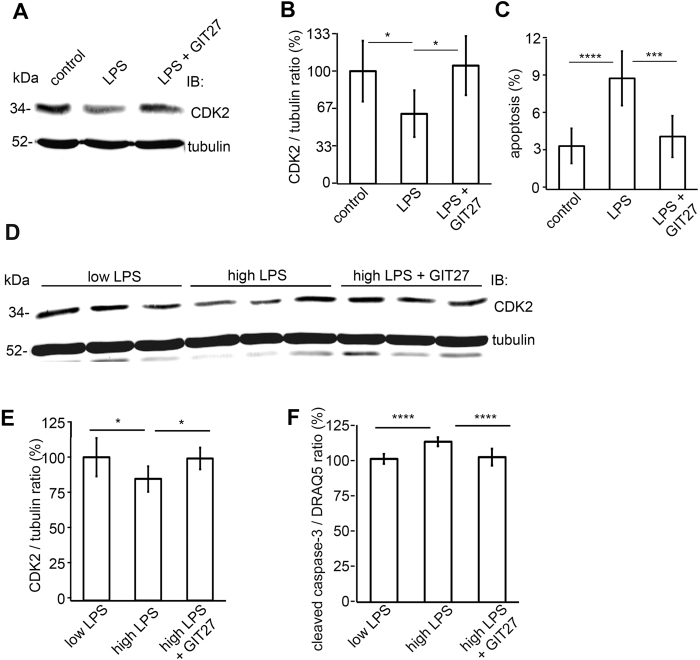
References
-
- Borch-Johnsen K. et al. The natural history of insulin-dependent diabetes mellitus in Denmark: 1. Long-term survival with and without late diabetic complications. Diabet. Med. 4, 201–210 (1987). - PubMed
-
- Reddy G. R., Kotlyarevska K., Ransom R. F. & Menon R. K. The podocyte and diabetes mellitus: is the podocyte the key to the origins of diabetic nephropathy? Curr. Opin. Nephrol. Hypertens. 17, 32–36 (2008). - PubMed
-
- Steffes M. W., Schmidt D., McCrery R. & Basgen J. M. & International Diabetic Nephropathy Study Group. Glomerular cell number in normal subjects and in type 1 diabetic patients. Kidney Int. 59, 2104–2113 (2001). - PubMed
-
- Verzola D. et al. Apoptosis in the kidneys of patients with type II diabetic nephropathy. Kidney Int. 72, 1262–1272 (2007). - PubMed
-
- Meyer T. W., Bennett P. H. & Nelson R. G. Podocyte number predicts long-term urinary albumin excretion in Pima Indians with Type II diabetes and microalbuminuria. Diabetologia 42, 1341–1344 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
