

Heme oxygenase-1 inhibits basophil maturation and activation but promotes its apoptosis in T helper type 2-mediated allergic airway inflammation
- PMID: 26879758
- PMCID: PMC4754607
- DOI: 10.1111/imm.12564
Heme oxygenase-1 inhibits basophil maturation and activation but promotes its apoptosis in T helper type 2-mediated allergic airway inflammation
Abstract
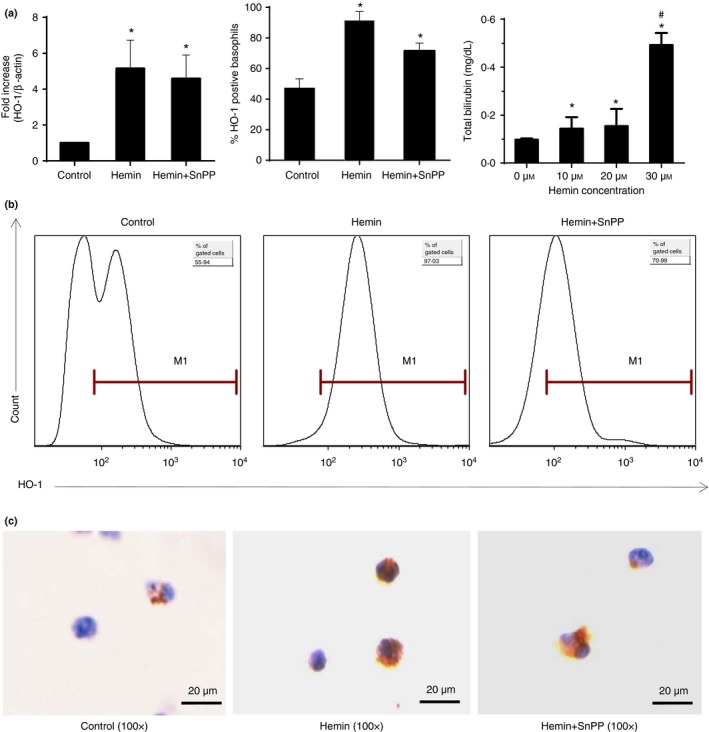
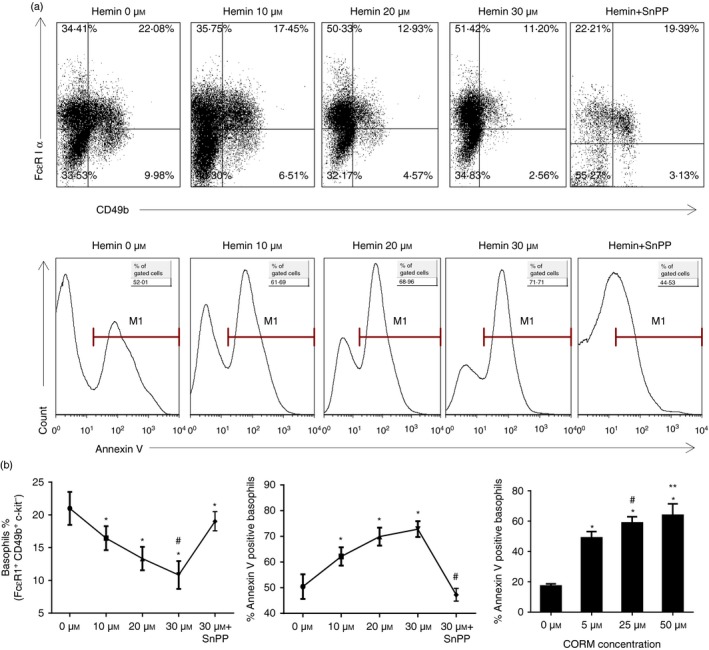
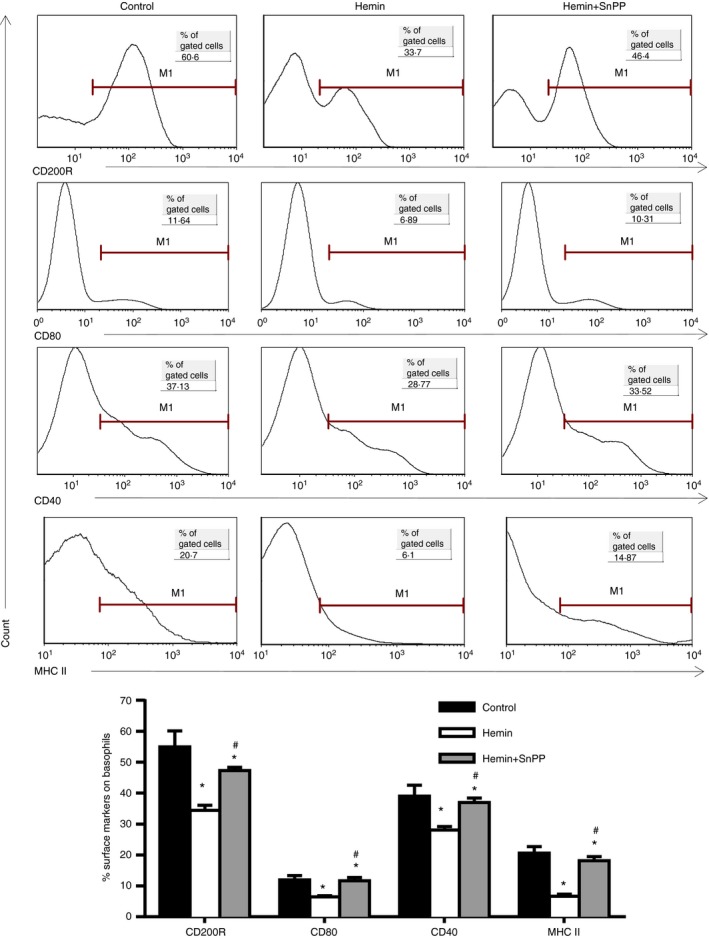
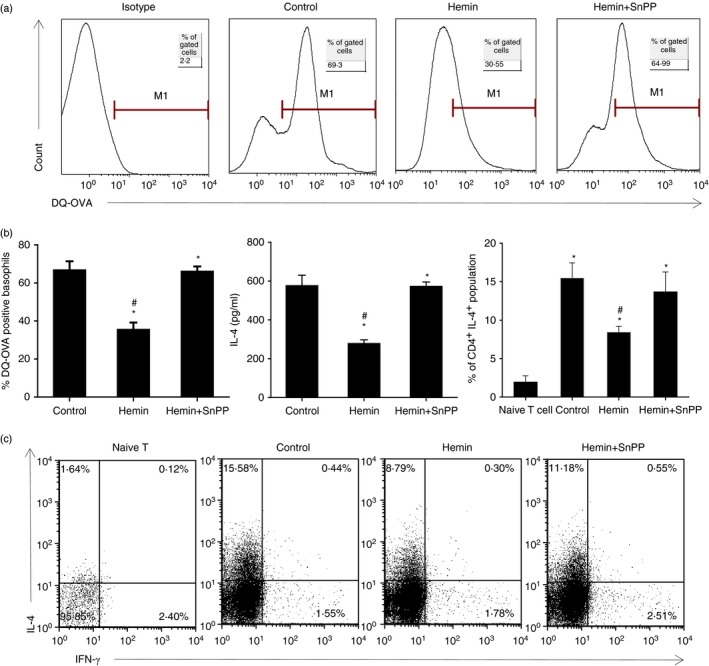
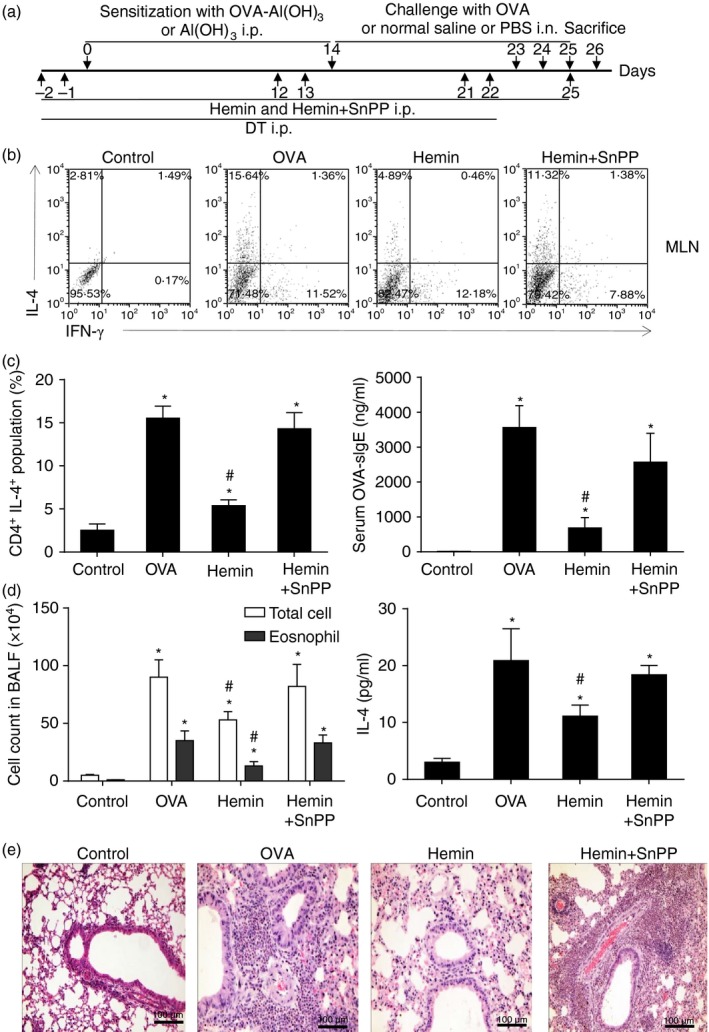
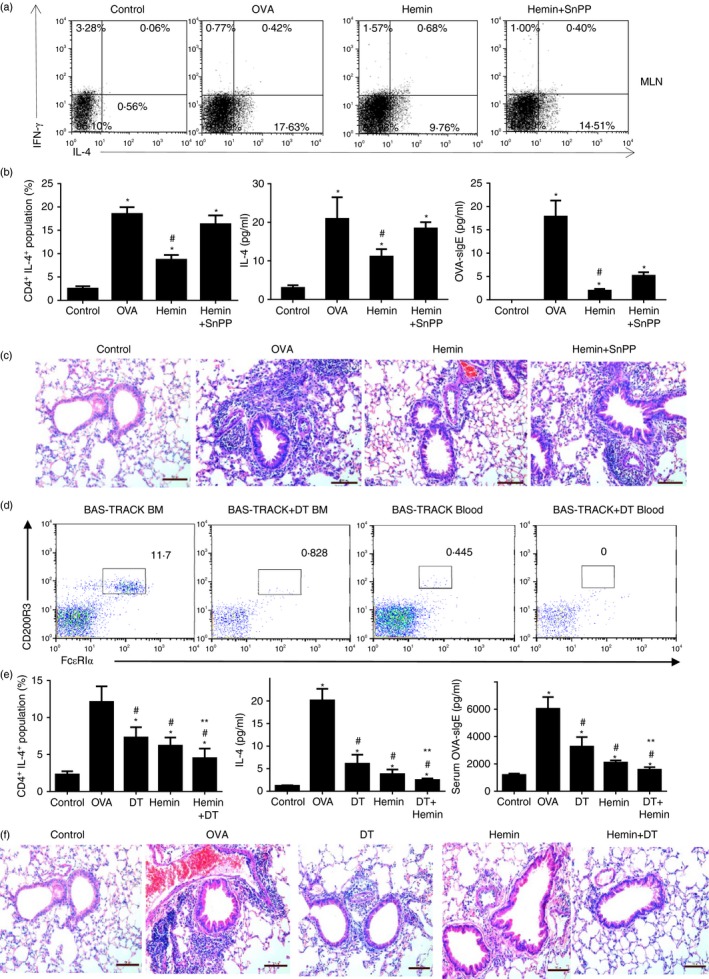
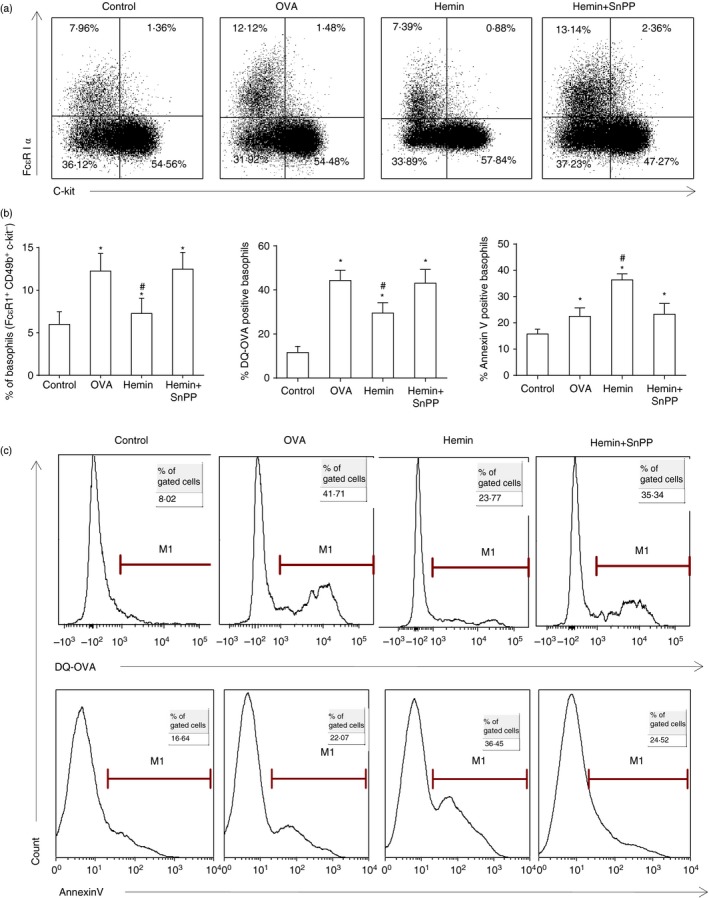
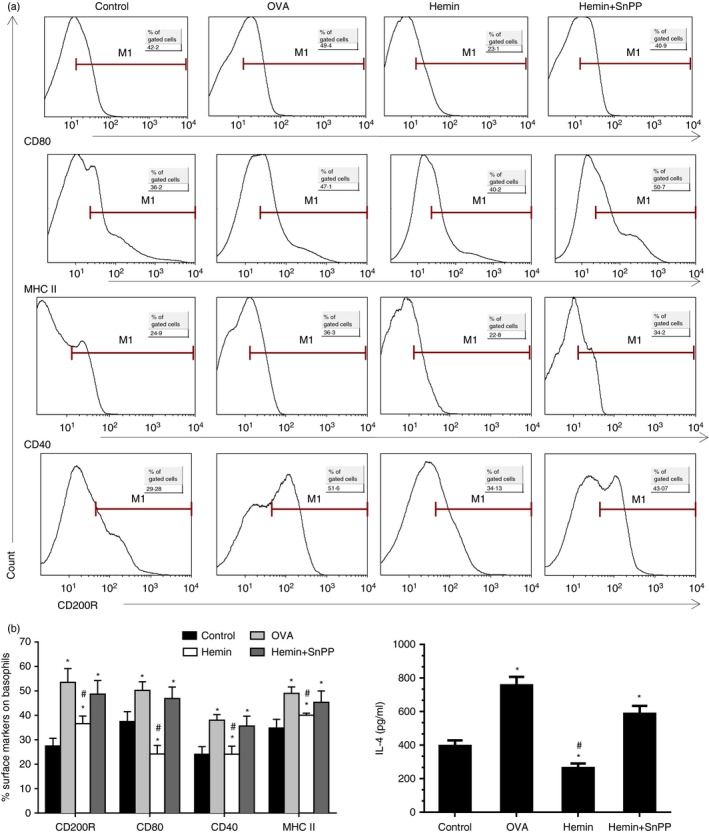
The anti-inflammatory role of heme oxygenase-1 (HO-1) has been studied extensively in many disease models including asthma. Many cell types are anti-inflammatory targets of HO-1, such as dendritic cells and regulatory T cells. In contrast to previous reports that HO-1 had limited effects on basophils, which participate in T helper type 2 immune responses and antigen-induced allergic airway inflammation, we demonstrated in this study, for the first time, that the up-regulation of HO-1 significantly suppressed the maturation of mouse basophils, decreased the expression of CD40, CD80, MHC-II and activation marker CD200R on basophils, blocked DQ-ovalbumin uptake and promoted basophil apoptosis both in vitro and in vivo, leading to the inhibition of T helper type 2 polarization. These effects of HO-1 were mimicked by exogenous carbon monoxide, which is one of the catalytic products of HO-1. Furthermore, adoptive transfer of HO-1-modified basophils reduced ovalbumin-induced allergic airway inflammation. The above effects of HO-1 can be reversed by the HO-1 inhibitor Sn-protoporphyrin IX. Moreover, conditional depletion of basophils accompanying hemin treatment further attenuated airway inflammation compared with the hemin group, indicating that the protective role of HO-1 may involve multiple immune cells. Collectively, our findings demonstrated that HO-1 exerted its anti-inflammatory function through suppression of basophil maturation and activation, but promotion of basophil apoptosis, providing a possible novel therapeutic target in allergic asthma.
Keywords: T helper type 2 response; allergic airway inflammation; basophil; heme oxygenase-1.
© 2015 John Wiley & Sons Ltd.
Figures
Similar articles
-
Basophils as a primary inducer of the T helper type 2 immunity in ovalbumin-induced allergic airway inflammation.Immunology. 2014 Jun;142(2):202-15. doi: 10.1111/imm.12240. Immunology. 2014. PMID: 24383680 Free PMC article.
-
Heme oxygenase-1 alleviates eosinophilic inflammation by inhibiting STAT3-SOCS3 signaling.Pediatr Pulmonol. 2020 Jun;55(6):1440-1447. doi: 10.1002/ppul.24759. Epub 2020 Apr 16. Pediatr Pulmonol. 2020. PMID: 32297710
-
Schisandrin B promotes Foxp3+ regulatory T cell expansion by activating heme oxygenase-1 in dendritic cells and exhibits immunomodulatory effects in Th2-mediated allergic asthma.Eur J Pharmacol. 2022 Mar 5;918:174775. doi: 10.1016/j.ejphar.2022.174775. Epub 2022 Jan 25. Eur J Pharmacol. 2022. PMID: 35085518
-
Recent advancement in the mechanism of basophil activation.J Dermatol Sci. 2018 Jul;91(1):3-8. doi: 10.1016/j.jdermsci.2018.03.007. Epub 2018 Mar 13. J Dermatol Sci. 2018. PMID: 29602578 Review.
-
Regulation of type 2 immunity by basophils.Adv Exp Med Biol. 2013;785:37-41. doi: 10.1007/978-1-4614-6217-0_4. Adv Exp Med Biol. 2013. PMID: 23456835 Review.
Cited by
-
Heme Catabolic Pathway in Inflammation and Immune Disorders.Front Pharmacol. 2019 Jul 24;10:825. doi: 10.3389/fphar.2019.00825. eCollection 2019. Front Pharmacol. 2019. PMID: 31396090 Free PMC article. Review.
-
Regional variations in allergen-induced airway inflammation correspond to changes in soluble guanylyl cyclase heme and expression of heme oxygenase-1.FASEB J. 2024 Mar 31;38(6):e23572. doi: 10.1096/fj.202301626RRR. FASEB J. 2024. PMID: 38512139 Free PMC article.
-
Airway epithelial TSLP production of TLR2 drives type 2 immunity in allergic airway inflammation.Eur J Immunol. 2018 Nov;48(11):1838-1850. doi: 10.1002/eji.201847663. Epub 2018 Oct 12. Eur J Immunol. 2018. PMID: 30184256 Free PMC article.
-
The Pharmacological Effect of Hemin in Inflammatory-Related Diseases: Protocol for a Systematic Review.JMIR Res Protoc. 2023 Nov 16;12:e48368. doi: 10.2196/48368. JMIR Res Protoc. 2023. PMID: 37971806 Free PMC article.
-
Utility of basophil activation test for predicting the outcome of wheezing in children: a pilot study.BMC Immunol. 2021 Jan 6;22(1):4. doi: 10.1186/s12865-020-00395-4. BMC Immunol. 2021. PMID: 33407109 Free PMC article.
References
-
- Naito Y, Takagi T, Higashimura Y. Heme oxygenase‐1 and anti‐inflammatory M2 macrophages. Arch Biochem Biophys 2014; 564C:83–8. - PubMed
-
- Gayathri G, Muthukumar S, Joseph LD, Suresh R. Immunolocalization of heme oxygenase‐1 in periodontal diseases. Indian J Dent Res 2014; 25:567–71. - PubMed
-
- Xia ZW, Zhong WW, Meyrowitz JS, Zhang ZL. The role of heme oxygenase‐1 in T cell‐mediated immunity: the all encompassing enzyme. Curr Pharm Des 2008; 14:454–64. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
