

MRN, CtIP, and BRCA1 mediate repair of topoisomerase II-DNA adducts
- PMID: 26880199
- PMCID: PMC4754713
- DOI: 10.1083/jcb.201504005
MRN, CtIP, and BRCA1 mediate repair of topoisomerase II-DNA adducts
Abstract
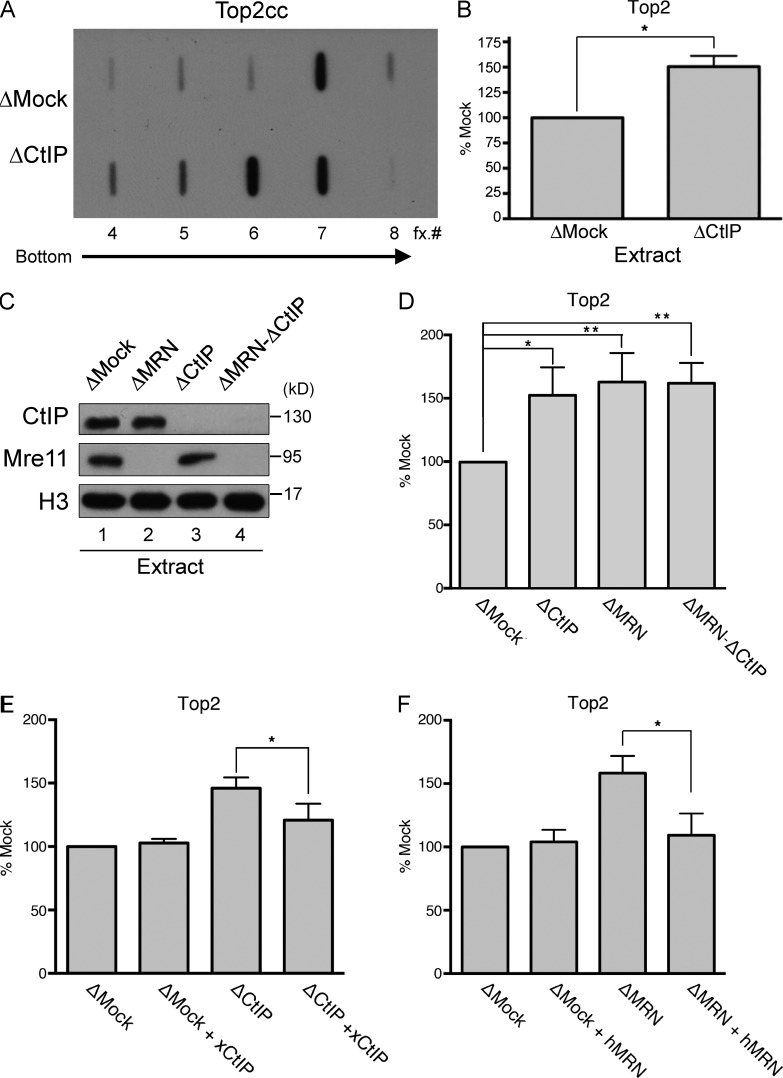
Repair of DNA double-strand breaks (DSBs) with complex ends poses a special challenge, as additional processing is required before DNA ligation. For example, protein-DNA adducts must be removed to allow repair by either nonhomologous end joining or homology-directed repair. Here, we investigated the processing of topoisomerase II (Top2)-DNA adducts induced by treatment with the chemotherapeutic agent etoposide. Through biochemical analysis in Xenopus laevis egg extracts, we establish that the MRN (Mre11, Rad50, and Nbs1) complex, CtIP, and BRCA1 are required for both the removal of Top2-DNA adducts and the subsequent resection of Top2-adducted DSB ends. Moreover, the interaction between CtIP and BRCA1, although dispensable for resection of endonuclease-generated DSB ends, is required for resection of Top2-adducted DSBs, as well as for cellular resistance to etoposide during genomic DNA replication.
© 2016 Aparicio et al.
Figures





References
-
- Bunting S.F., Callén E., Kozak M.L., Kim J.M., Wong N., López-Contreras A.J., Ludwig T., Baer R., Faryabi R.B., Malhowski A., et al. 2012. BRCA1 functions independently of homologous recombination in DNA interstrand crosslink repair. Mol. Cell. 46:125–135. 10.1016/j.molcel.2012.02.015 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
