

Myeloid-Derived Suppressor Cells Express Bruton's Tyrosine Kinase and Can Be Depleted in Tumor-Bearing Hosts by Ibrutinib Treatment
- PMID: 26880800
- PMCID: PMC4873459
- DOI: 10.1158/0008-5472.CAN-15-1490
Myeloid-Derived Suppressor Cells Express Bruton's Tyrosine Kinase and Can Be Depleted in Tumor-Bearing Hosts by Ibrutinib Treatment
Abstract
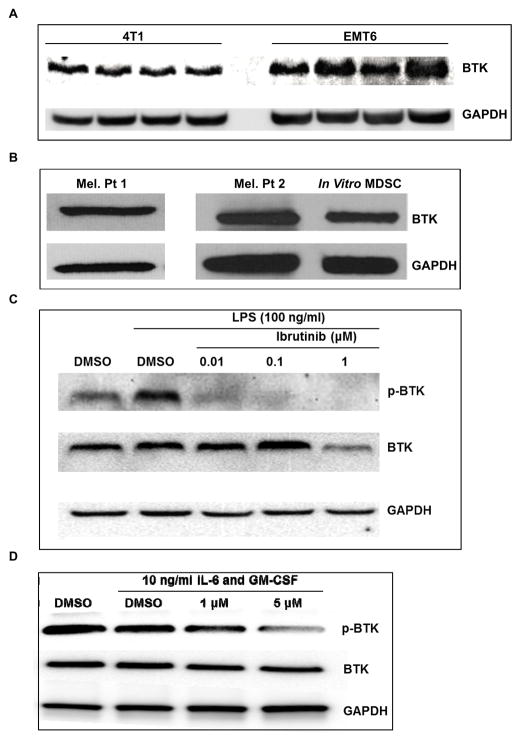
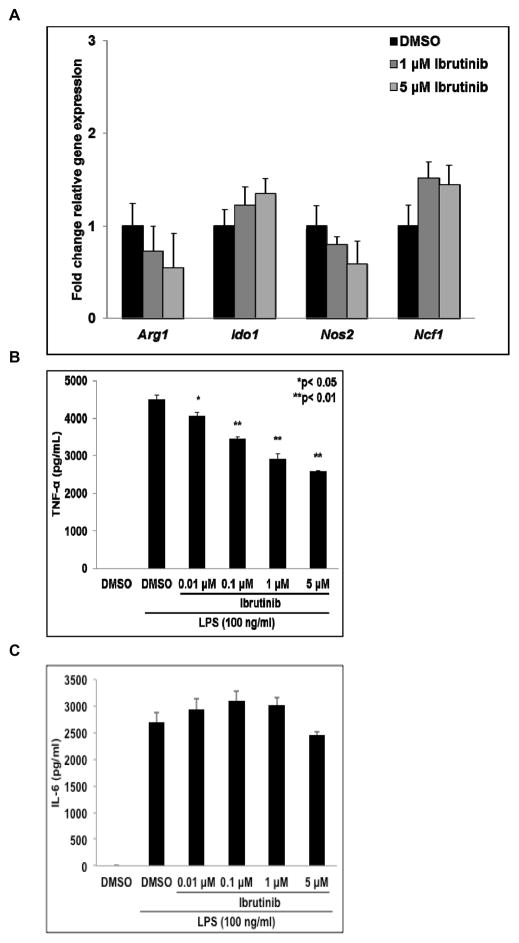
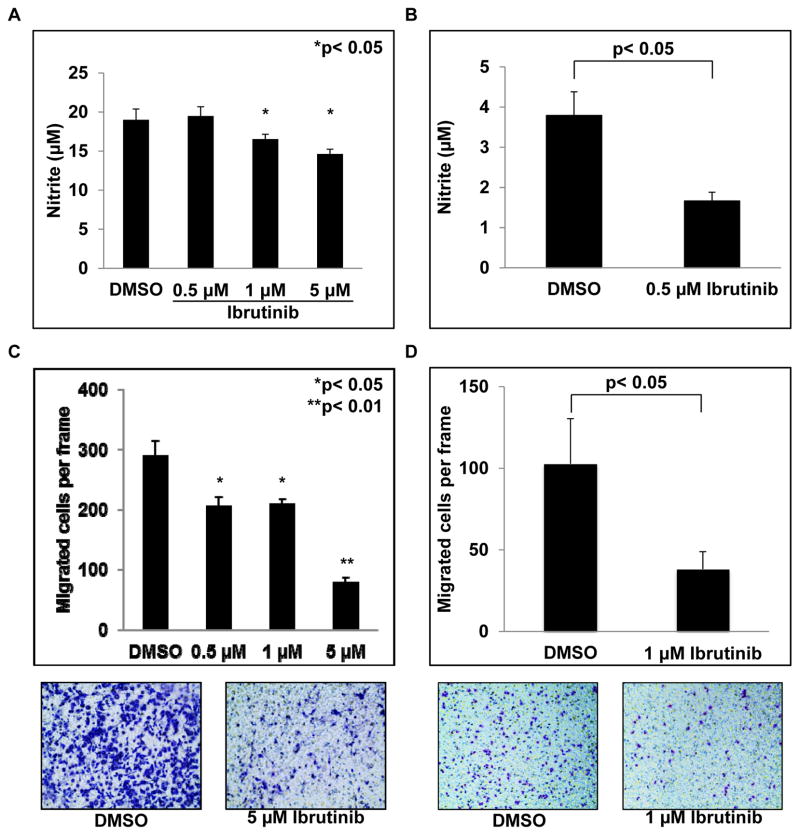
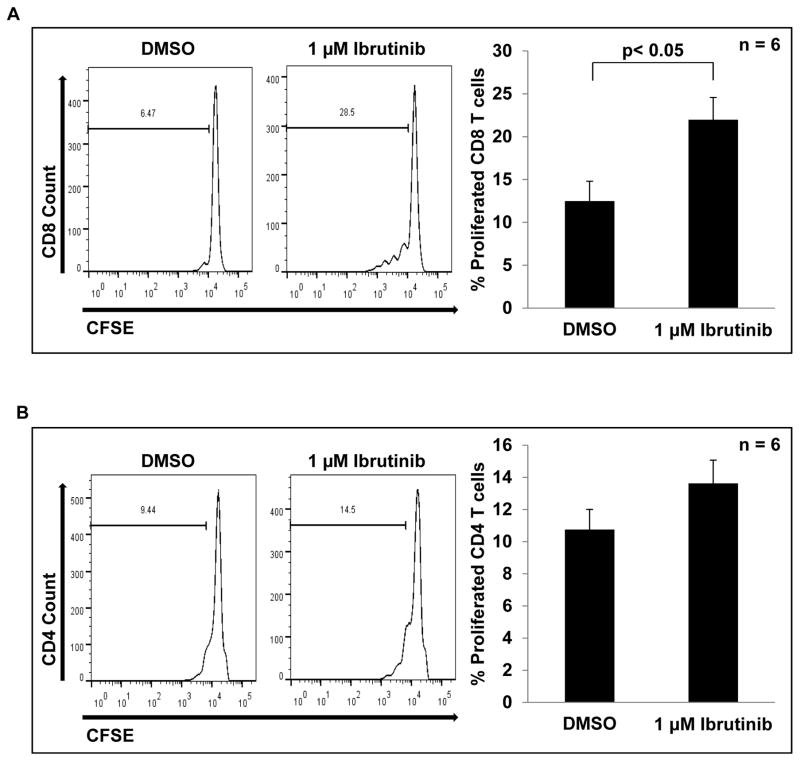
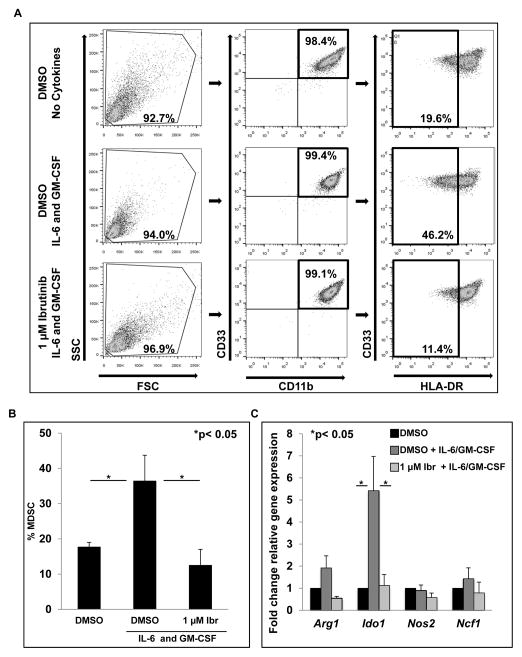
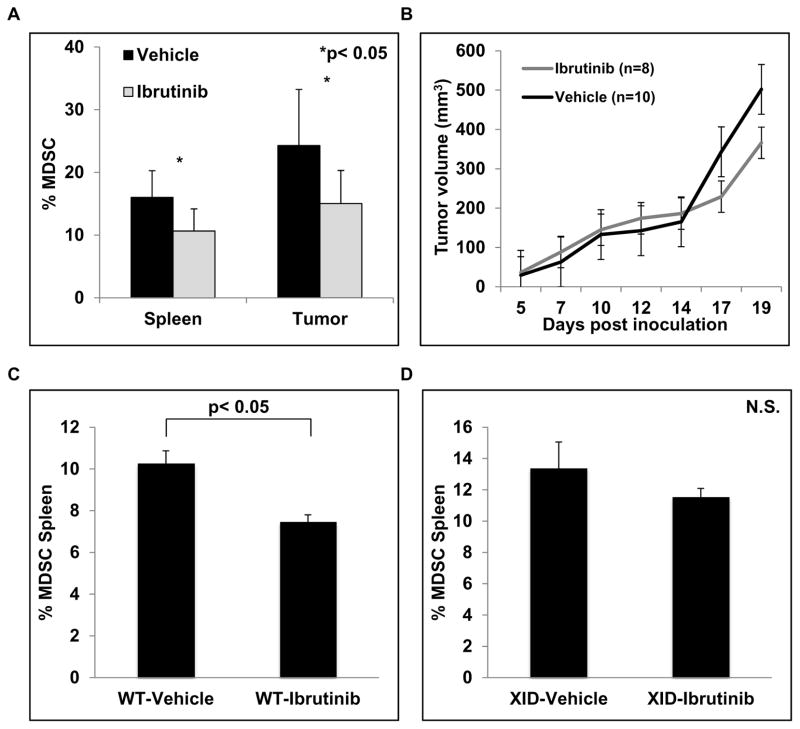
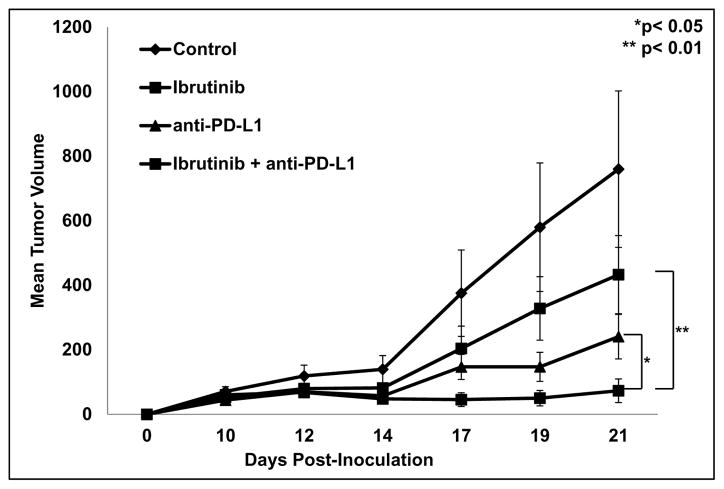
Myeloid-derived suppressor cells (MDSC) are a heterogeneous group of immature myeloid cells that expand in tumor-bearing hosts in response to soluble factors produced by tumor and stromal cells. MDSC expansion has been linked to loss of immune effector cell function and reduced efficacy of immune-based cancer therapies, highlighting the MDSC population as an attractive therapeutic target. Ibrutinib, an irreversible inhibitor of Bruton's tyrosine kinase (BTK) and IL2-inducible T-cell kinase (ITK), is in clinical use for the treatment of B-cell malignancies. Here, we report that BTK is expressed by murine and human MDSCs, and that ibrutinib is able to inhibit BTK phosphorylation in these cells. Treatment of MDSCs with ibrutinib significantly impaired nitric oxide production and cell migration. In addition, ibrutinib inhibited in vitro generation of human MDSCs and reduced mRNA expression of indolamine 2,3-dioxygenase, an immunosuppressive factor. Treatment of mice bearing EMT6 mammary tumors with ibrutinib resulted in reduced frequency of MDSCs in both the spleen and tumor. Ibrutinib treatment also resulted in a significant reduction of MDSCs in wild-type mice bearing B16F10 melanoma tumors, but not in X-linked immunodeficiency mice (XID) harboring a BTK mutation, suggesting that BTK inhibition plays an important role in the observed reduction of MDSCs in vivo Finally, ibrutinib significantly enhanced the efficacy of anti-PD-L1 (CD274) therapy in a murine breast cancer model. Together, these results demonstrate that ibrutinib modulates MDSC function and generation, revealing a potential strategy for enhancing immune-based therapies in solid malignancies. Cancer Res; 76(8); 2125-36. ©2016 AACR.
©2016 American Association for Cancer Research.
Conflict of interest statement
The authors have no conflicts of interest to disclose.
Figures
References
-
- Melani C, Chiodoni C, Forni G, Colombo MP. Myeloid cell expansion elicited by the progression of spontaneous mammary carcinomas in c-erbB-2 transgenic BALB/c mice suppresses immune reactivity. Blood. 2003;102(6):2138–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
