

Stability properties of neuronal microtubules
- PMID: 26887570
- PMCID: PMC5541393
- DOI: 10.1002/cm.21286
Stability properties of neuronal microtubules
Abstract
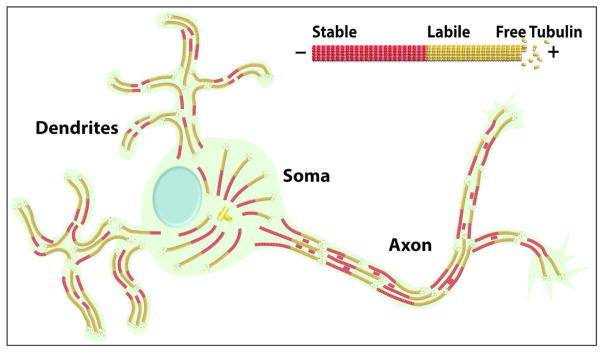
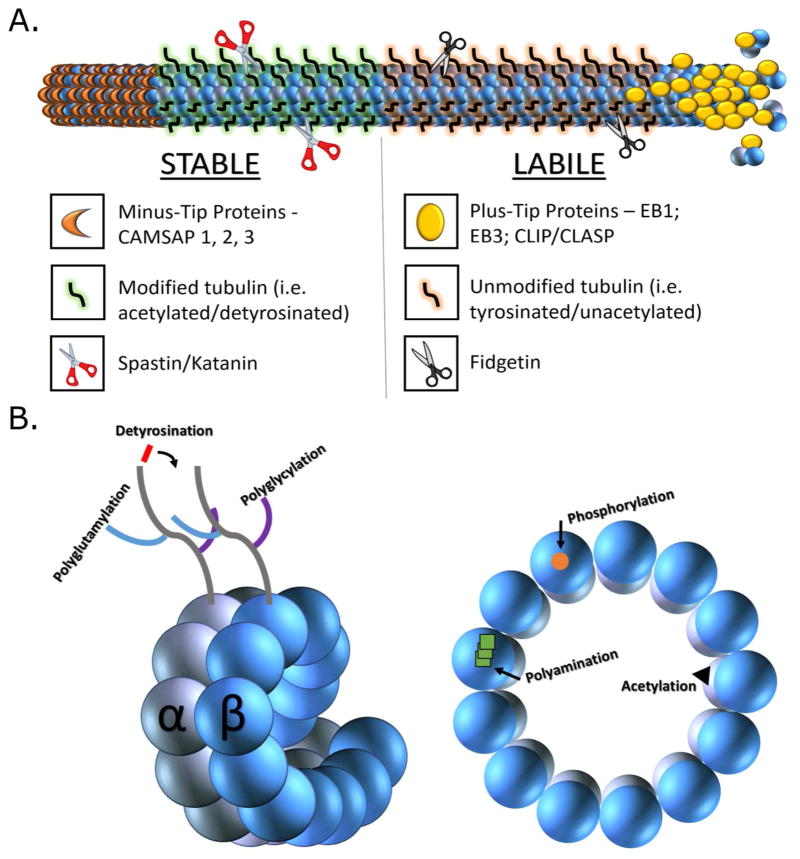
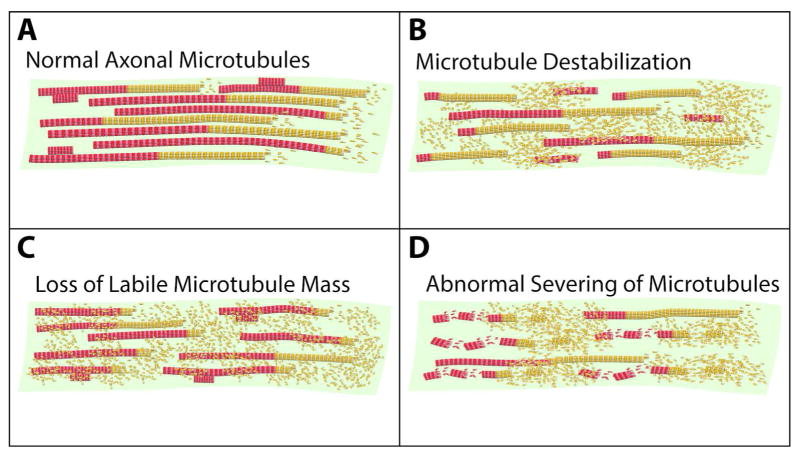
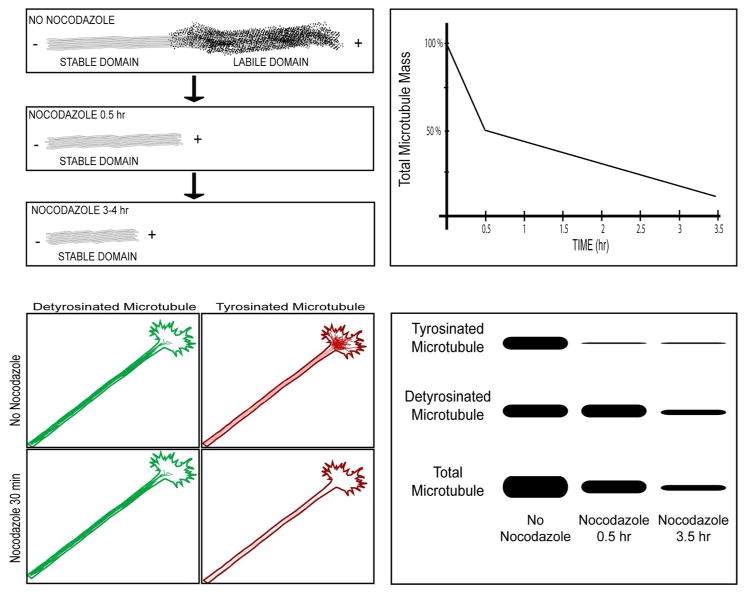
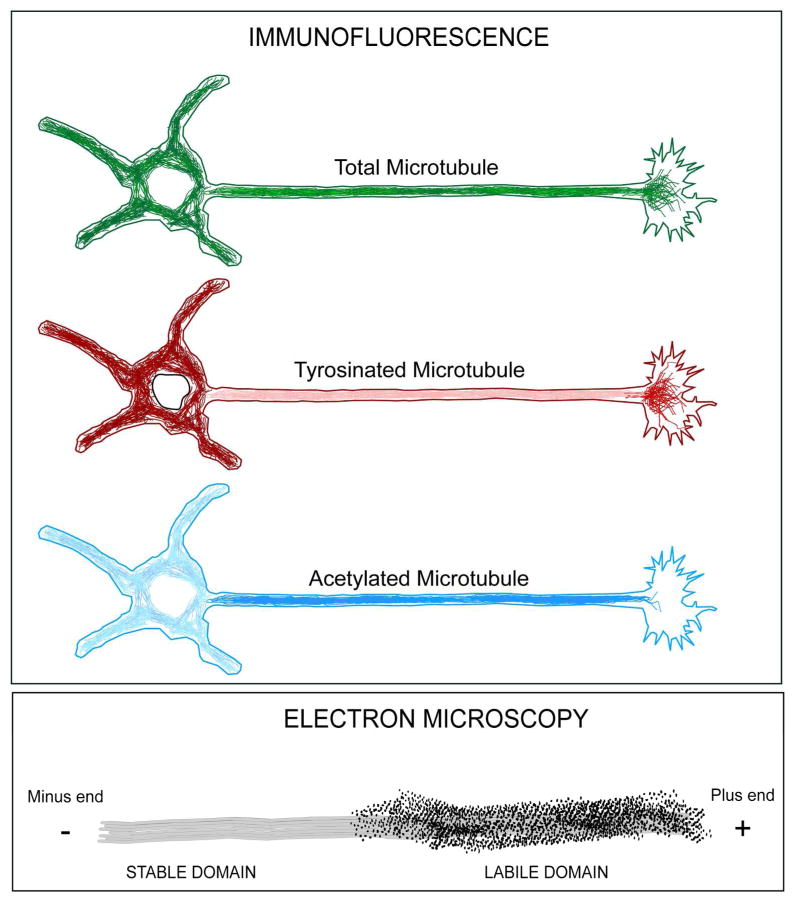
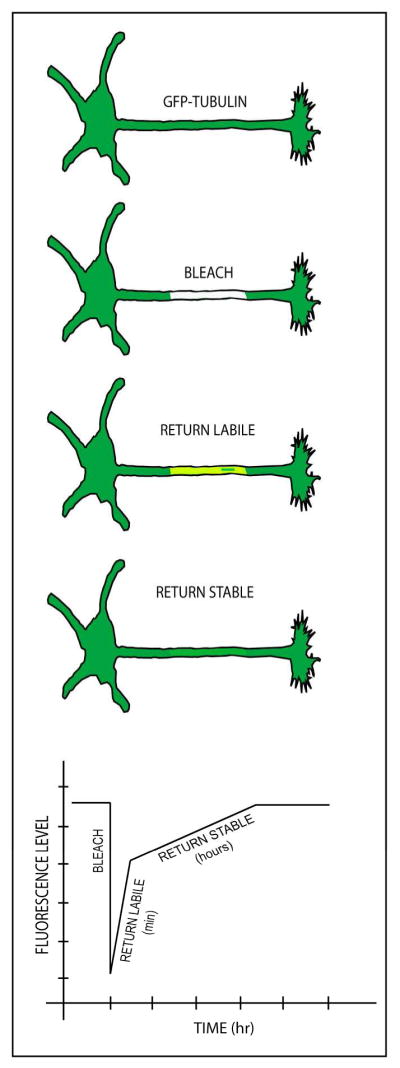
Neurons are terminally differentiated cells that use their microtubule arrays not for cell division but rather as architectural elements required for the elaboration of elongated axons and dendrites. In addition to acting as compression-bearing struts that provide for the shape of the neuron, microtubules also act as directional railways for organelle transport. The stability properties of neuronal microtubules are commonly discussed in the biomedical literature as crucial to the development and maintenance of the nervous system, and have recently gained attention as central to the etiology of neurodegenerative diseases. Drugs that affect microtubule stability are currently under investigation as potential therapies for disease and injury of the nervous system. There is often a lack of consistency, however, in how the issue of microtubule stability is discussed in the literature, and this can affect the design and interpretation of experiments as well as potential therapeutic regimens. Neuronal microtubules are considered to be more stable than microtubules in dividing cells. On average, this is true, but in addition to an abundant stable microtubule fraction in neurons, there is also an abundant labile microtubule fraction. Both are functionally important. Individual microtubules consist of domains that differ in their stability properties, and these domains can also differ markedly in their composition as well as how they interact with various microtubule-related proteins in the neuron. Myriad proteins and pathways have been discussed as potential contributors to microtubule stability in neurons. © 2016 Wiley Periodicals, Inc.
Keywords: +tip; Alzheimer's disease; CAMSAP; MAP6; acetylation; axon; dendrite; detyrosination; fidgetin; katanin; labile; microtubule; microtubule stability; neurodegeneration; neuron; nocodazole; polyamination; spastin; stable; tau; taxol; tubulin.
© 2016 Wiley Periodicals, Inc.
Figures
References
-
- Ahmad FJ, Baas PW. Microtubules released from the neuronal centrosome are transported into the axon. Journal of cell science. 1995;108(Pt 8):2761–2769. - PubMed
-
- Ahmad FJ, Joshi HC, Centonze VE, Baas PW. Inhibition of microtubule nucleation at the neuronal centrosome compromises axon growth. Neuron. 1994;12:271–280. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
