

Concise Review: Control of Cell Fate Through Cell Cycle and Pluripotency Networks
- PMID: 26889666
- PMCID: PMC5201256
- DOI: 10.1002/stem.2345
Concise Review: Control of Cell Fate Through Cell Cycle and Pluripotency Networks
Abstract
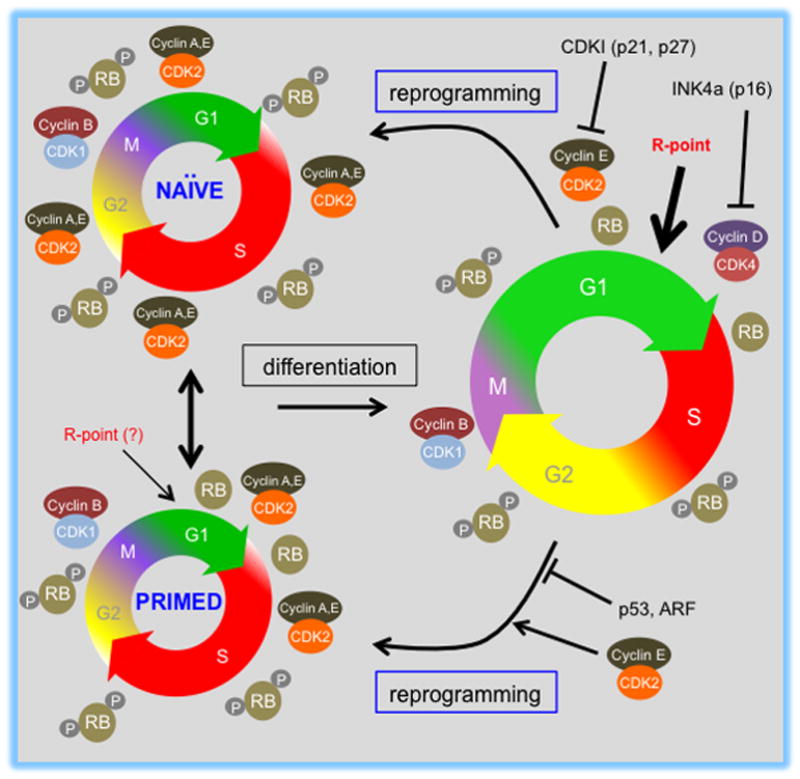
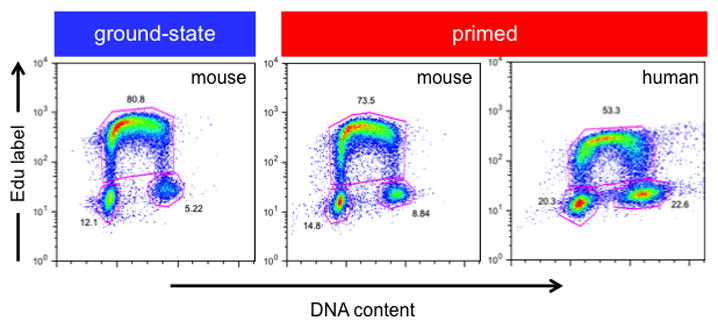
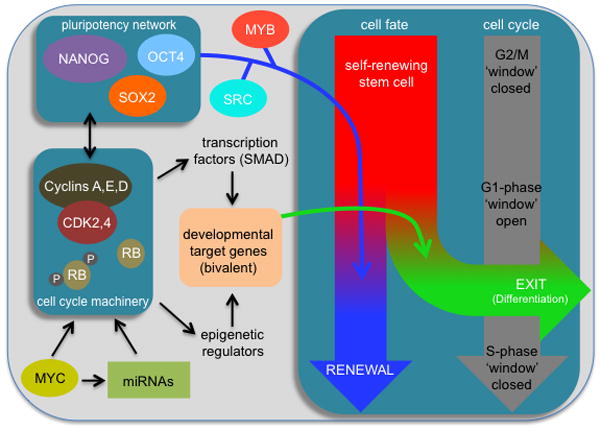
Pluripotent stem cells (PSCs) proliferate rapidly with a characteristic cell cycle structure consisting of short G1- and G2-gap phases. This applies broadly to PSCs of peri-implantation stage embryos, cultures of embryonic stem cells, induced pluripotent stem cells, and embryonal carcinoma cells. During the early stages of PSC differentiation however, cell division times increase as a consequence of cell cycle remodeling. Most notably, this is indicated by elongation of the G1-phase. Observations linking changes in the cell cycle with exit from pluripotency have raised questions about the role of cell cycle control in maintenance of the pluripotent state. Until recently however, this has been a difficult question to address because of limitations associated with experimental tools. Recent studies now show that pluripotency and cell cycle regulatory networks are intertwined and that cell cycle control mechanisms are an integral, mechanistic part of the PSC state. Studies in embryonal carcinoma, some 30 years ago, first suggested that pluripotent cells initiate differentiation when in the G1-phase. More recently, a molecular "priming" mechanism has been proposed to explain these observations in human embryonic stem cells. Complexity in this area has been increased by the realization that pluripotent cells exist in multiple developmental states and that in addition to each having their own characteristic gene expression and epigenetic signatures, they potentially have alternate modes of cell cycle regulation. This review will summarize current knowledge in these areas and will highlight important aspects of interconnections between the cell cycle, self-renewal, pluripotency, and cell fate decisions. Stem Cells 2016;34:1427-1436.
Keywords: Cell cycle; Differentiation; Embryonic stem cells; Pluripotent stem cells.
© 2016 AlphaMed Press.
Figures
References
-
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. CELL. 2006;126(4):663–676. - PubMed
-
- Heasman J. Patterning the early Xenopus embryo. DEVELOPMENT. 2006;133(7):1205–1217. - PubMed
-
- Yarden A, Salomon D, Geiger B. Zebrafish cyclin D1 is differentially expressed during early embryogenesis. BIOCHIM BIOPHYS ACTA. 1995;1264(3):257–260. - PubMed
-
- Kane DA, Kimmel CB. The zebrafish midblastula transition. DEVELOPMENT. 1993;119(2):447–456. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
