

Protective effects of butyrate-based compounds on a mouse model for spinal muscular atrophy
- PMID: 26892876
- PMCID: PMC4834225
- DOI: 10.1016/j.expneurol.2016.02.009
Protective effects of butyrate-based compounds on a mouse model for spinal muscular atrophy
Abstract
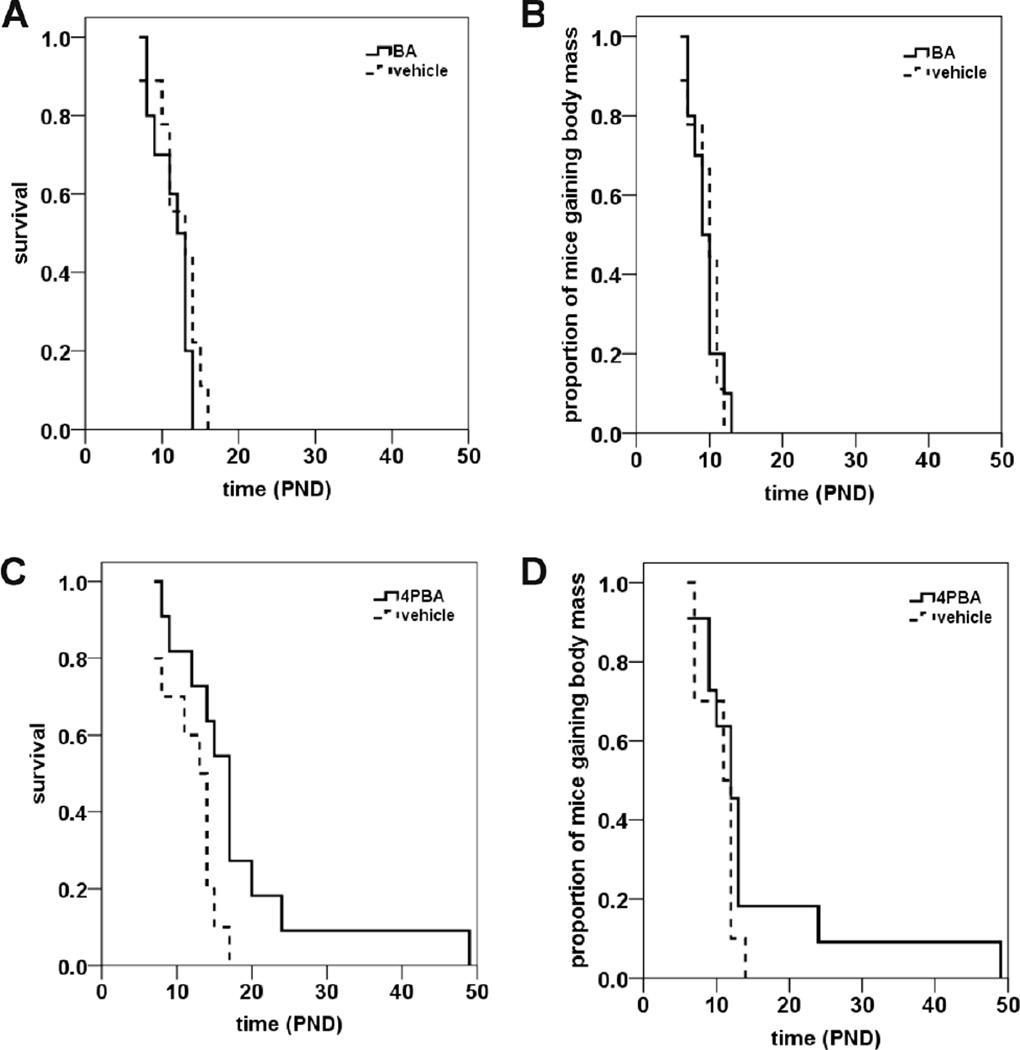
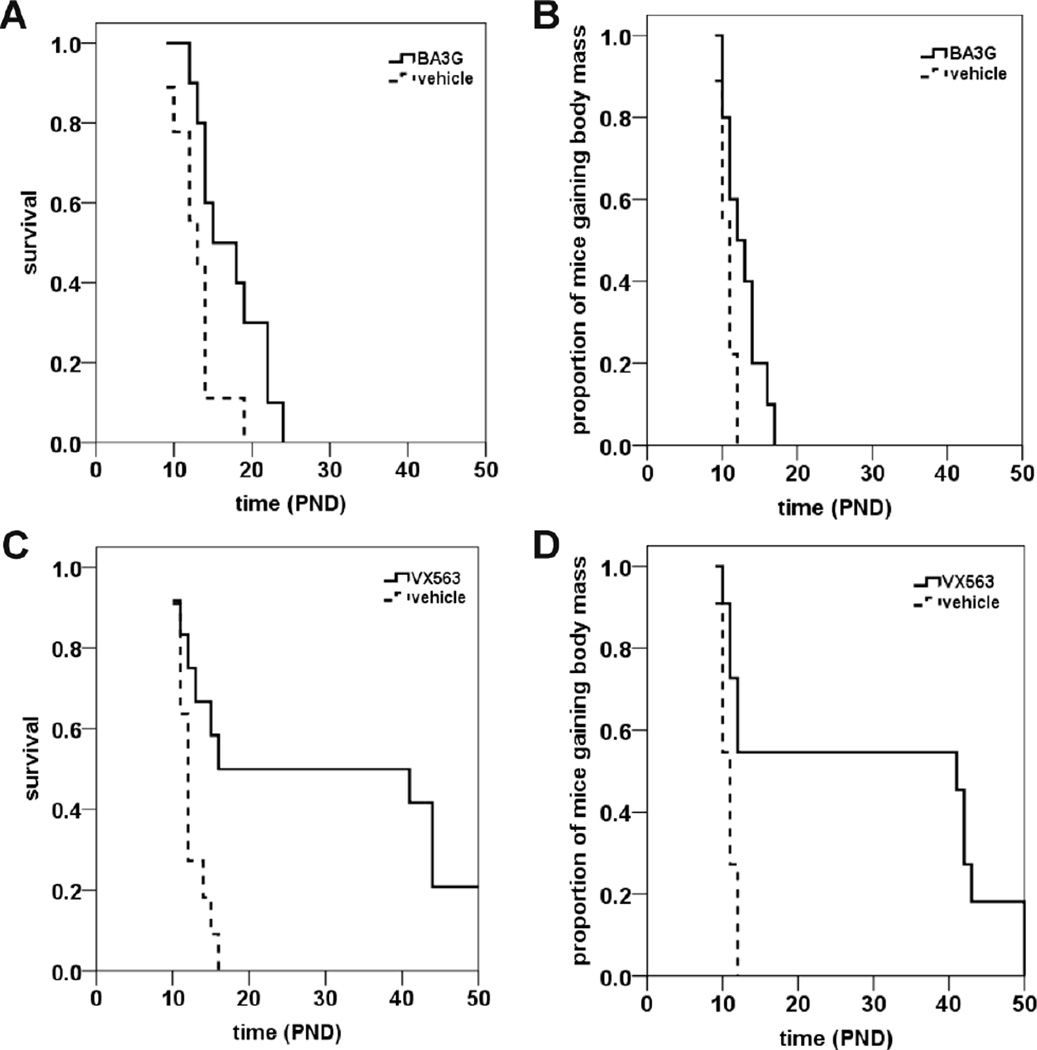
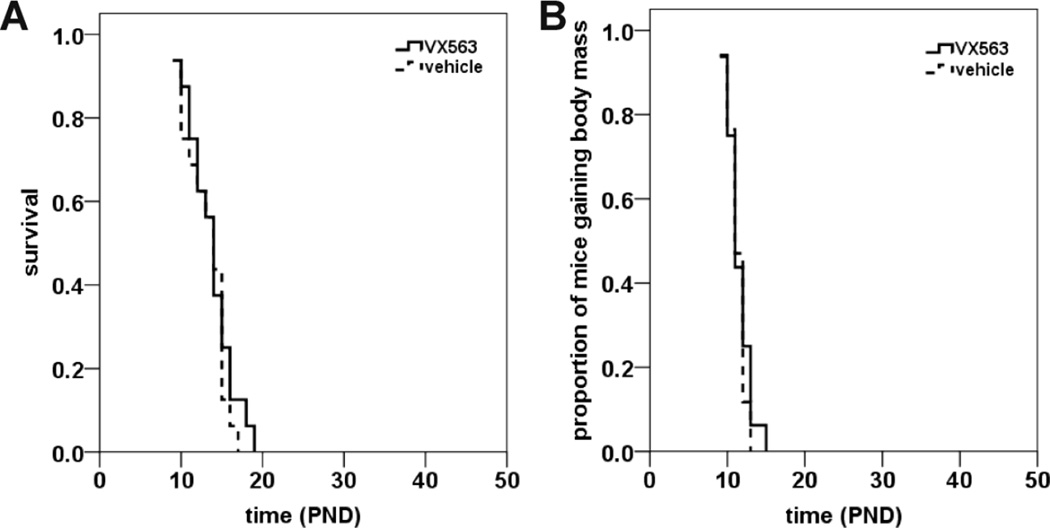
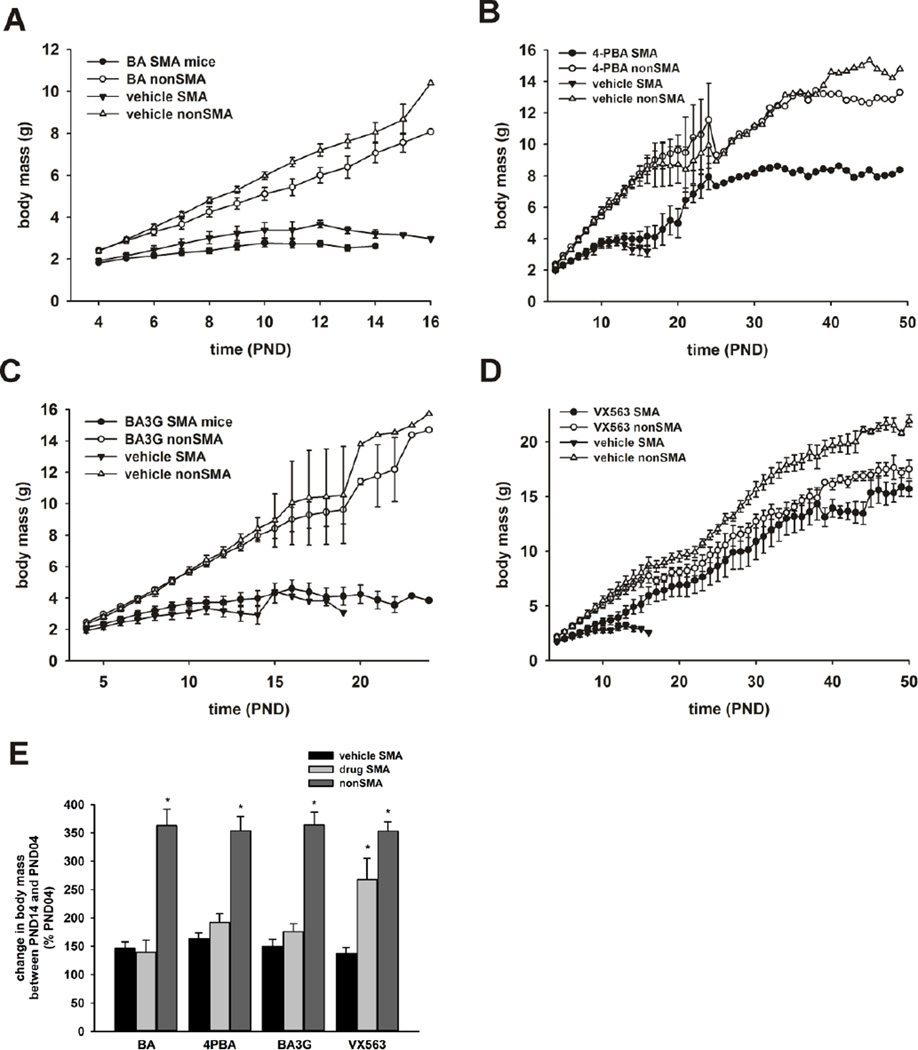
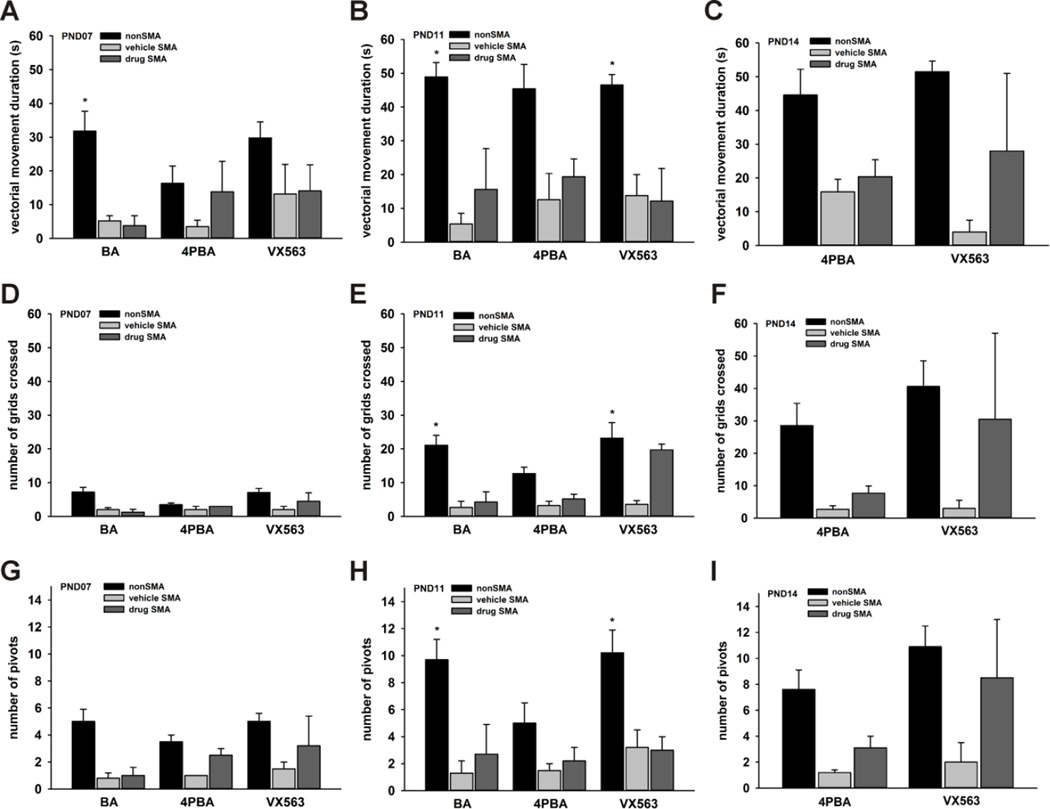
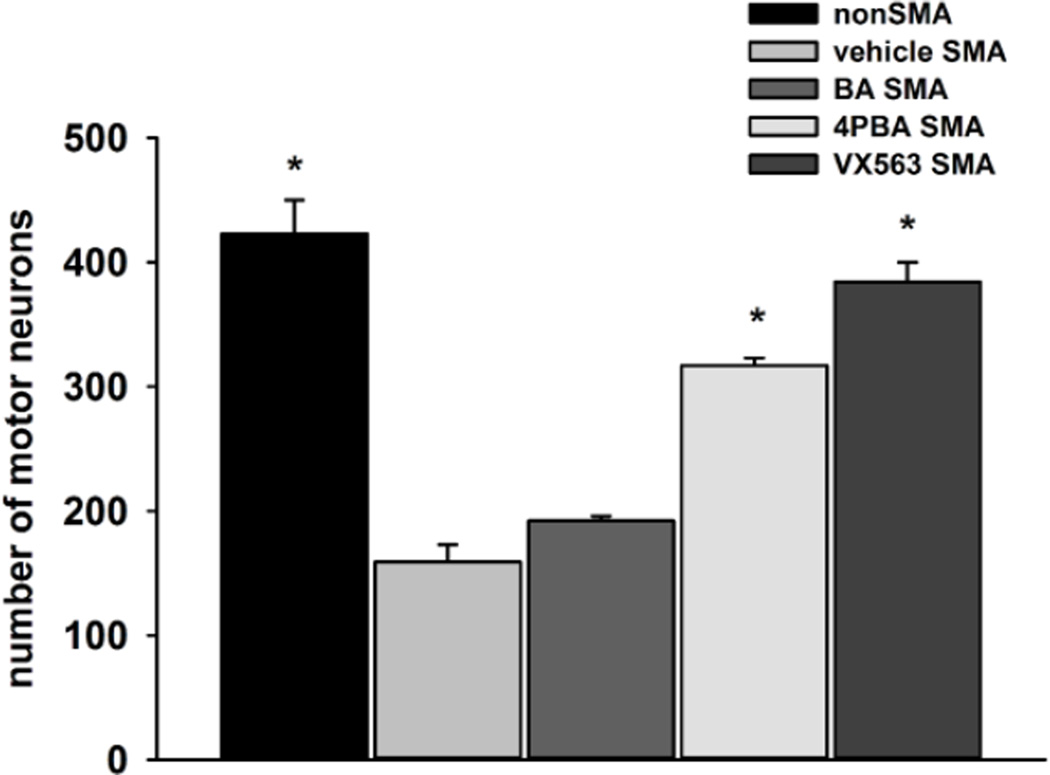
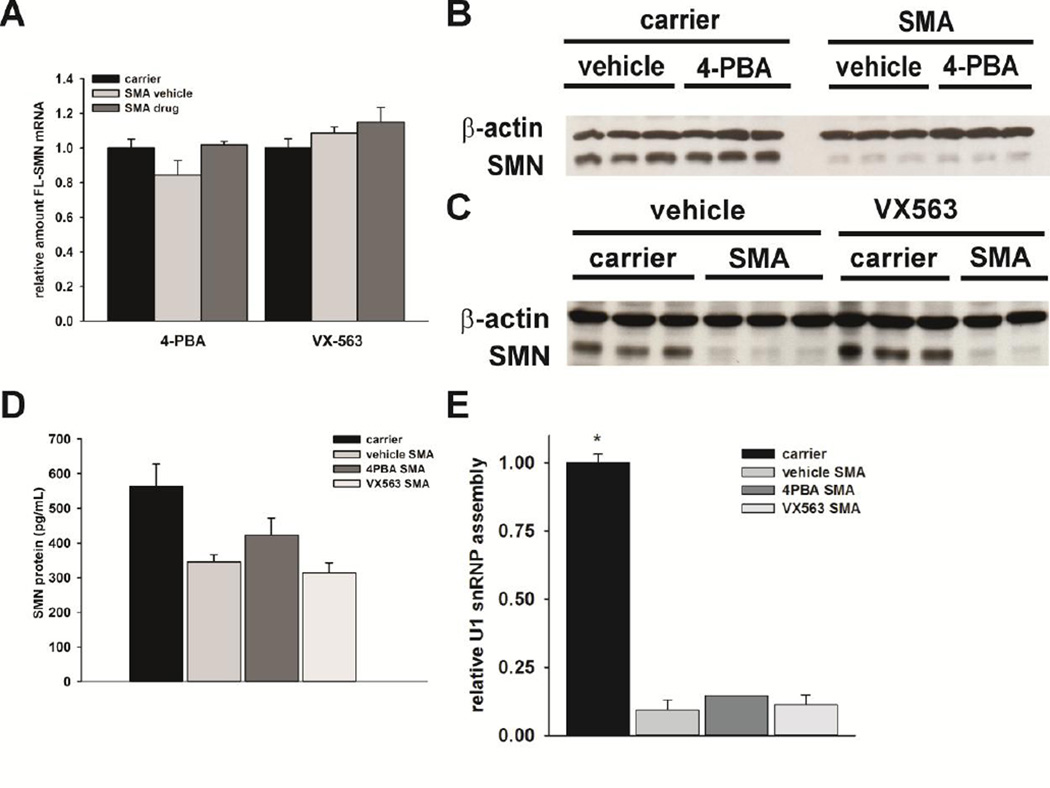
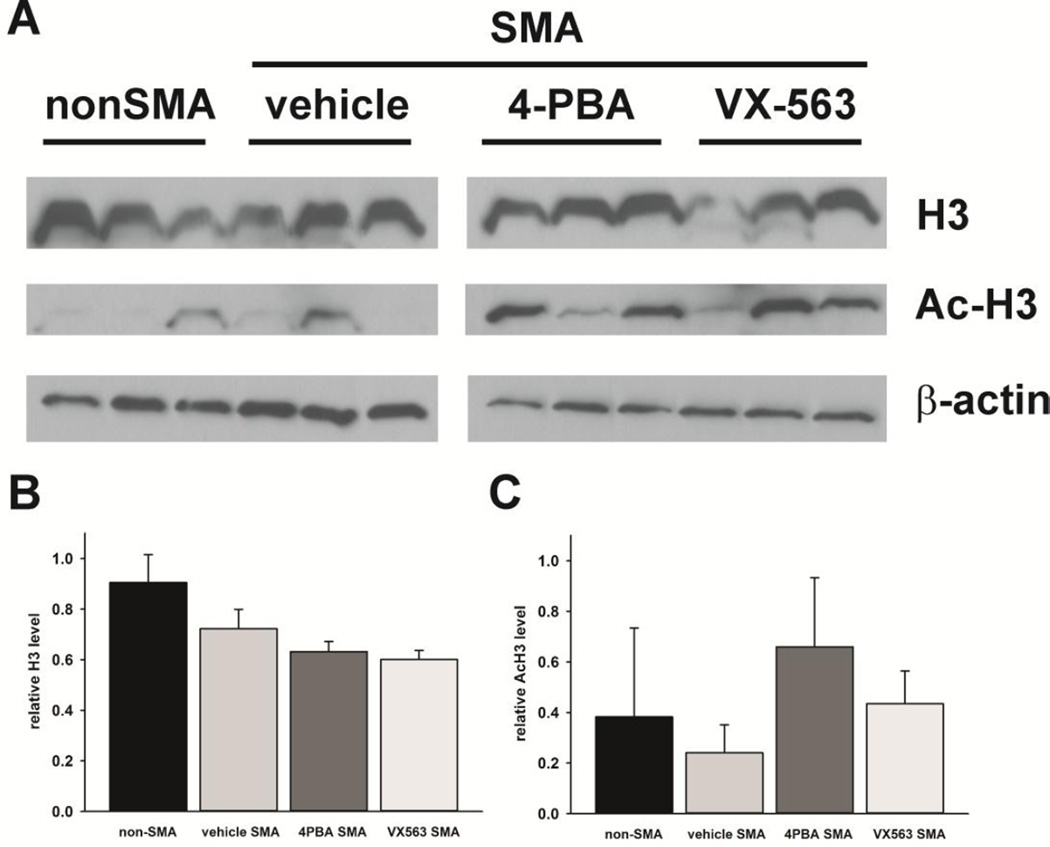
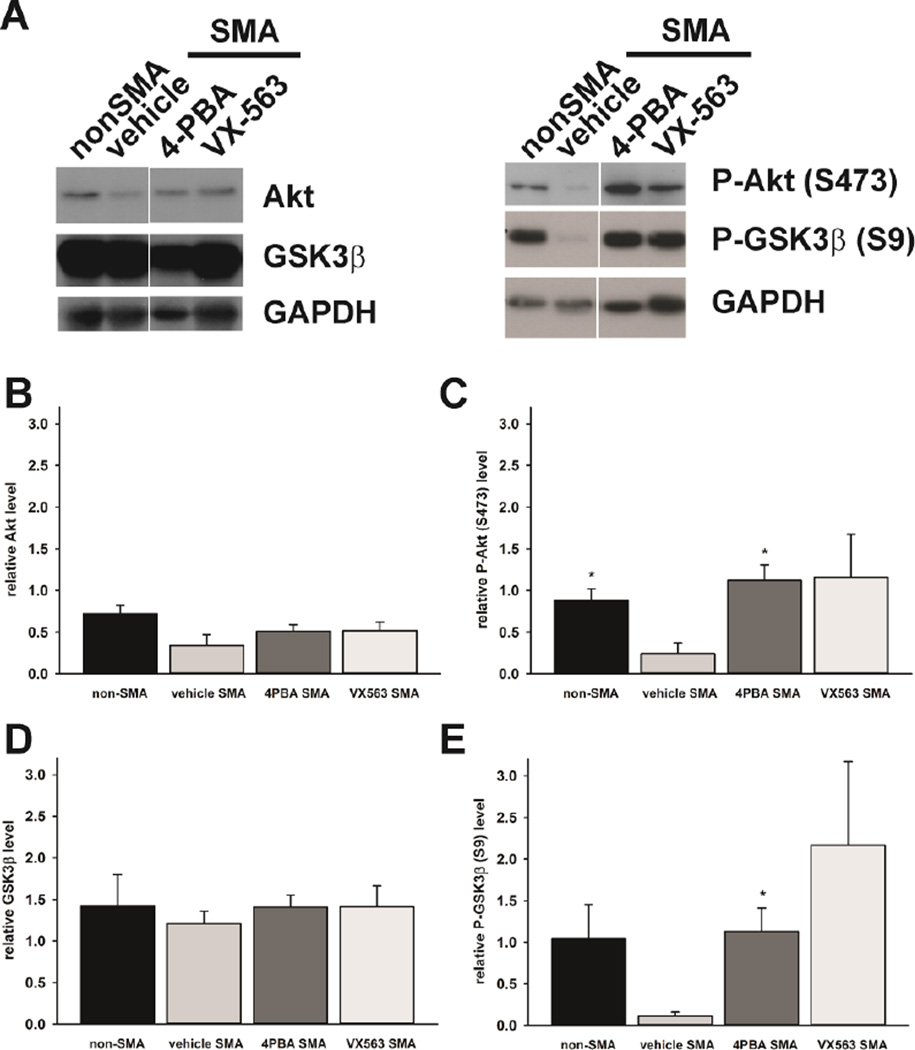
Proximal spinal muscular atrophy (SMA) is a childhood-onset degenerative disease resulting from the selective loss of motor neurons in the spinal cord. SMA is caused by the loss of SMN1 (survival motor neuron 1) but retention of SMN2. The number of copies of SMN2 modifies disease severity in SMA patients as well as in mouse models, making SMN2 a target for therapeutics development. Sodium butyrate (BA) and its analog (4PBA) have been shown to increase SMN2 expression in SMA cultured cells. In this study, we examined the effects of BA, 4PBA as well as two BA prodrugs-glyceryl tributyrate (BA3G) and VX563-on the phenotype of SMNΔ7 SMA mice. Treatment with 4PBA, BA3G and VX563 but not BA beginning at PND04 significantly improved the lifespan and delayed disease end stage, with administration of VX563 also improving the growth rate of these mice. 4PBA and VX563 improved the motor phenotype of SMNΔ7 SMA mice and prevented spinal motor neuron loss. Interestingly, neither 4PBA nor VX563 had an effect on SMN expression in the spinal cords of treated SMNΔ7 SMA mice; however, they inhibited histone deacetylase (HDAC) activity and restored the normal phosphorylation states of Akt and glycogen synthase kinase 3β, both of which are altered by SMN deficiency in vivo. These observations show that BA-based compounds with favorable pharmacokinetics ameliorate SMA pathology possibly by modulating HDAC and Akt signaling.
Keywords: 4-phenylbutyrate; Akt; Butyrate prodrug; Neuroprotection; Preclinical drug trial; Spinal muscular atrophy.
Copyright © 2016 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no significant conflicts of interest.
Figures
References
-
- Andreassi C, Angelozzi C, Tiziano FD, Vitali T, De Vincenzi E, Boninsegna A, Villanova M, Bertini E, Pini A, Neri G, Brahe C. Phenylbutyrate increases SMN expression in vitro: relevance for treatment of spinal muscular atrophy. Eur. J. Hum. Genet. 2004;12:59–65. - PubMed
-
- Berg S, Serabe B, Aleksic A, Bomgaars L, McGuffey L, Dauser R, Durfee J, Nuchtern J, Blaney S. Pharmacokinetics and cerebrospinal fluid penetration of phenylacetate and phenylbutyrate in the nonhuman primate. Cancer Chemother. Pharmacol. 2001;47:385–390. - PubMed
-
- Biondi O, Branchu J, Ben Salah A, Houdebine L, Bertin L, Chali F, Desseille C, Weill L, Sanchez G, Lancelin C, Aïd S, Lopes P, Pariset C, Lécolle S, Côté J, Holzenberger M, Chanoine G, Massaad C, Charbonnier F. IGF-1R reduction triggers neuroprotective signaling pathways in spinal muscular atrophy mice. J. Neurosci. 2015;35:12063–12079. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
