

Three-dimensional structures of two heavily N-glycosylated Aspergillus sp. family GH3 β-D-glucosidases
- PMID: 26894673
- PMCID: PMC4756609
- DOI: 10.1107/S2059798315024237
Three-dimensional structures of two heavily N-glycosylated Aspergillus sp. family GH3 β-D-glucosidases
Abstract
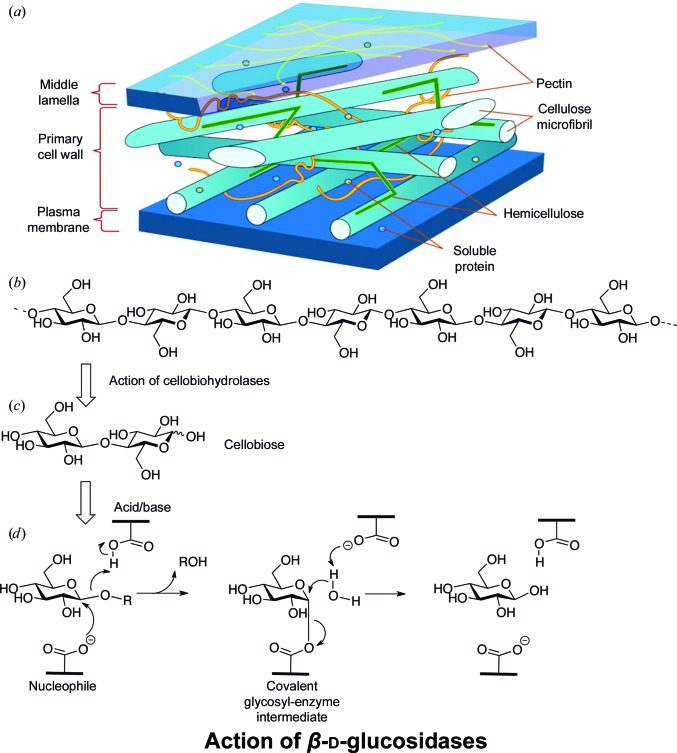
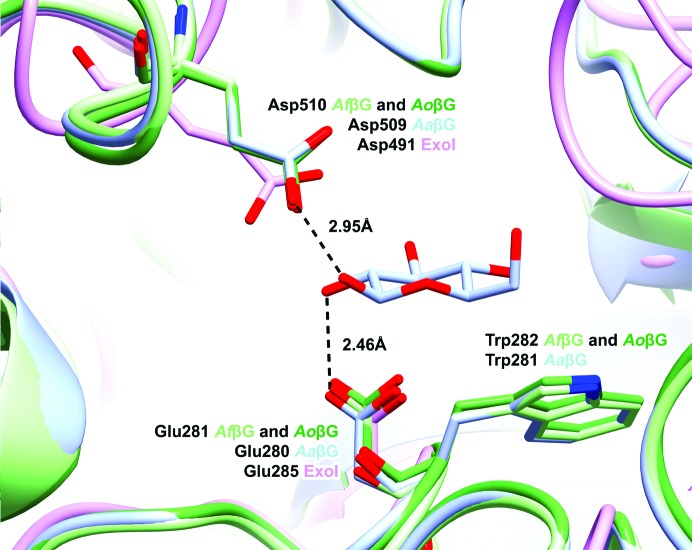
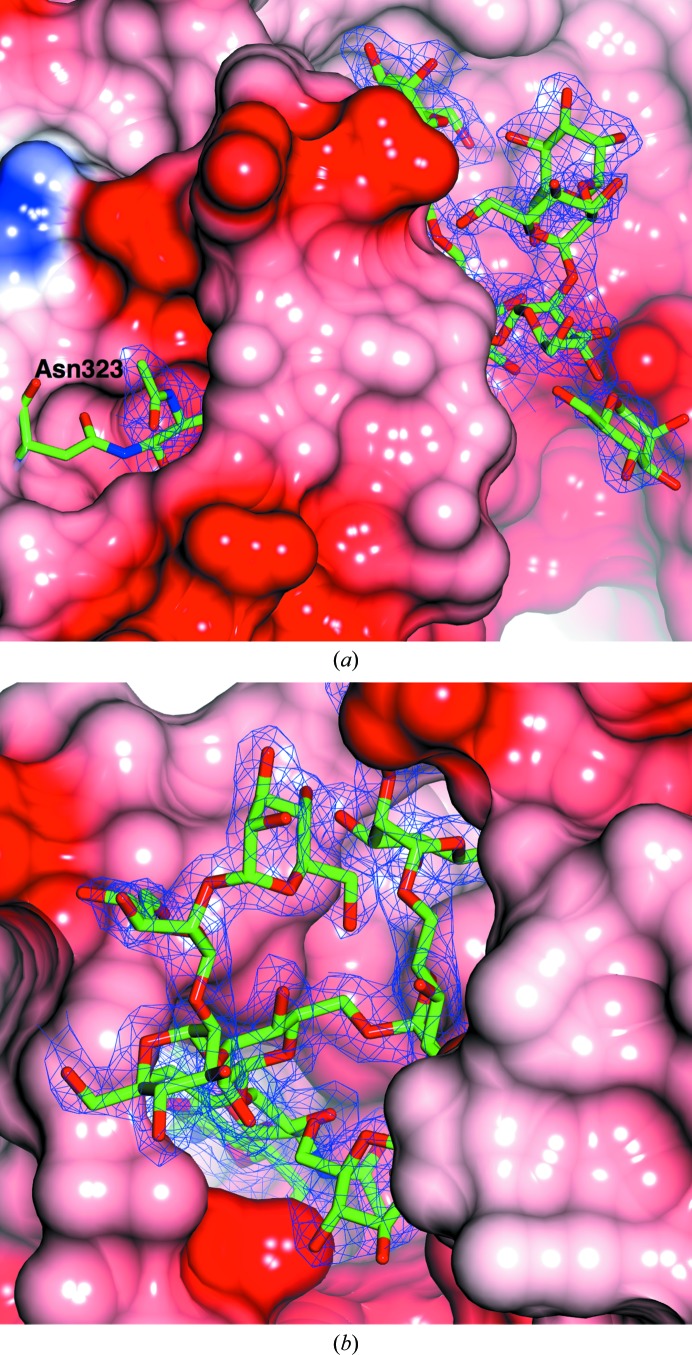
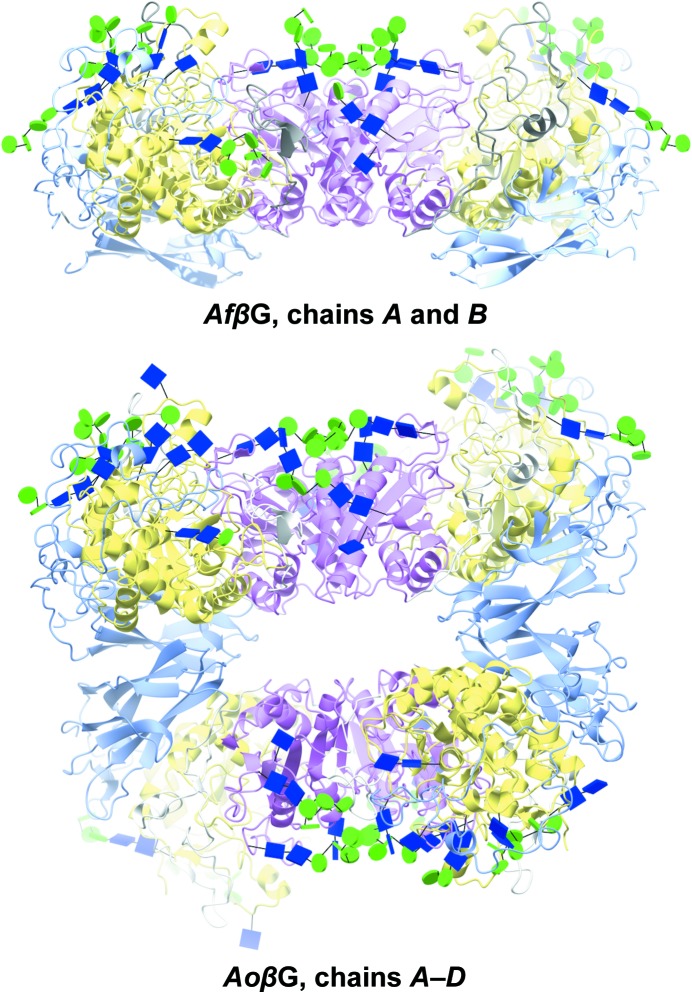
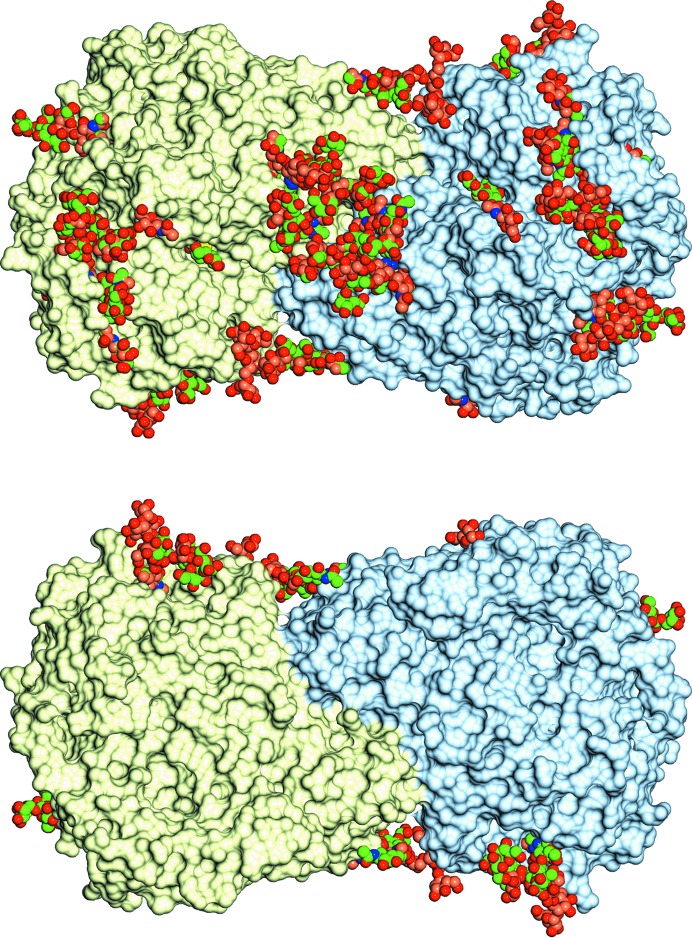
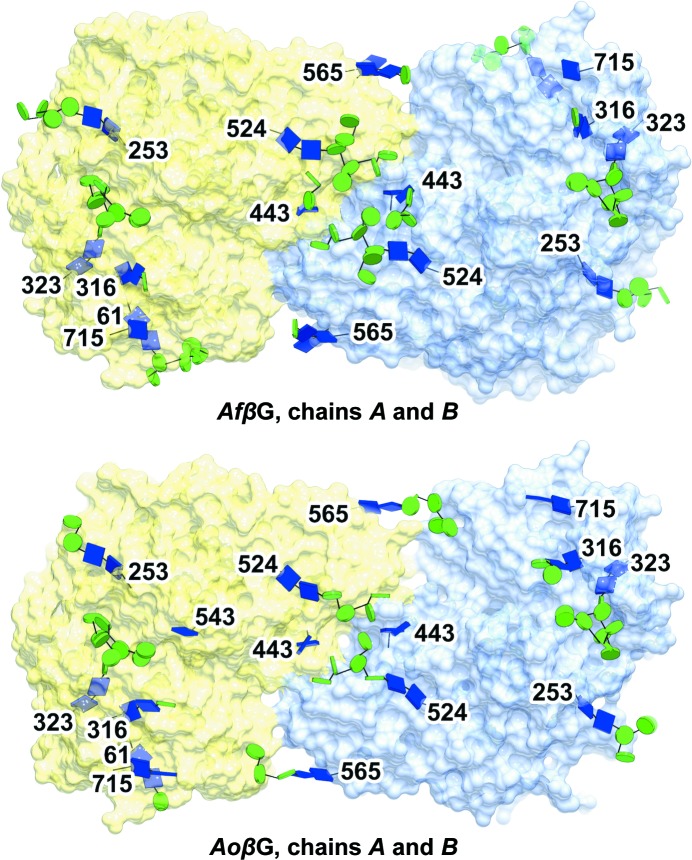
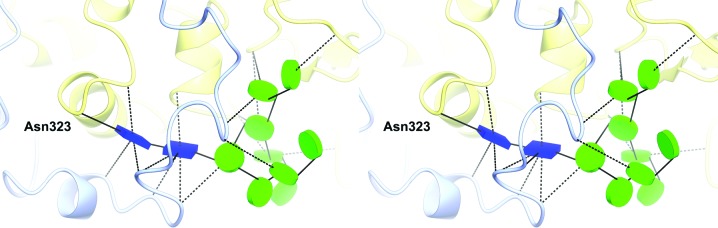
The industrial conversion of cellulosic plant biomass into useful products such as biofuels is a major societal goal. These technologies harness diverse plant degrading enzymes, classical exo- and endo-acting cellulases and, increasingly, cellulose-active lytic polysaccharide monooxygenases, to deconstruct the recalcitrant β-D-linked polysaccharide. A major drawback with this process is that the exo-acting cellobiohydrolases suffer from severe inhibition from their cellobiose product. β-D-Glucosidases are therefore important for liberating glucose from cellobiose and thereby relieving limiting product inhibition. Here, the three-dimensional structures of two industrially important family GH3 β-D-glucosidases from Aspergillus fumigatus and A. oryzae, solved by molecular replacement and refined at 1.95 Å resolution, are reported. Both enzymes, which share 78% sequence identity, display a three-domain structure with the catalytic domain at the interface, as originally shown for barley β-D-glucan exohydrolase, the first three-dimensional structure solved from glycoside hydrolase family GH3. Both enzymes show extensive N-glycosylation, with only a few external sites being truncated to a single GlcNAc molecule. Those glycans N-linked to the core of the structure are identified purely as high-mannose trees, and establish multiple hydrogen bonds between their sugar components and adjacent protein side chains. The extensive glycans pose special problems for crystallographic refinement, and new techniques and protocols were developed especially for this work. These protocols ensured that all of the D-pyranosides in the glycosylation trees were modelled in the preferred minimum-energy (4)C1 chair conformation and should be of general application to refinements of other crystal structures containing O- or N-glycosylation. The Aspergillus GH3 structures, in light of other recent three-dimensional structures, provide insight into fungal β-D-glucosidases and provide a platform on which to inform and inspire new generations of variant enzymes for industrial application.
Keywords: N-glycan; biofuels; cellulose degradation; glucosidase; glycoblocks.
Figures

References
-
- Agirre, J., Davies, G., Wilson, K. & Cowtan, K. (2015). Nature Chem. Biol. 11, 303. - PubMed
-
- Agirre, J., Iglesias-Fernandez, J., Rovira, C., Davies, G., Wilson, K. & Cowtan, K. (2015). Nature Struct. Mol. Biol. 22, 833–834. - PubMed
-
- Agrawal, R., Satlewal, A., Gaur, R., Mathur, A., Kumar, R., Gupta, R. P. & Tuli, D. K. (2015). Biochem. Eng. J. 102, 54–61.
-
- Ask, M., Olofsson, K., Di Felice, T., Ruohonen, L., Penttilä, M., Lidén, G. & Olsson, L. (2012). Process Biochem. 47, 1452–1459.
-
- Bacik, J.-P., Whitworth, G. E., Stubbs, K. A., Vocadlo, D. J. & Mark, B. L. (2012). Chem. Biol. 19, 1471–1482. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
