

Mechanisms of Bacterial (Serratia marcescens) Attachment to, Migration along, and Killing of Fungal Hyphae
- PMID: 26896140
- PMCID: PMC4836416
- DOI: 10.1128/AEM.04070-15
Mechanisms of Bacterial (Serratia marcescens) Attachment to, Migration along, and Killing of Fungal Hyphae
Abstract
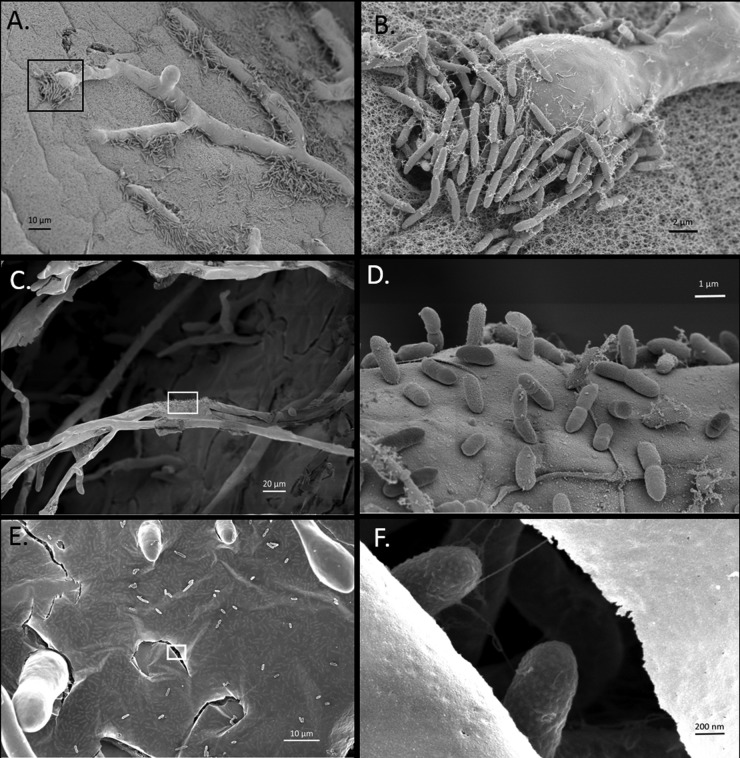
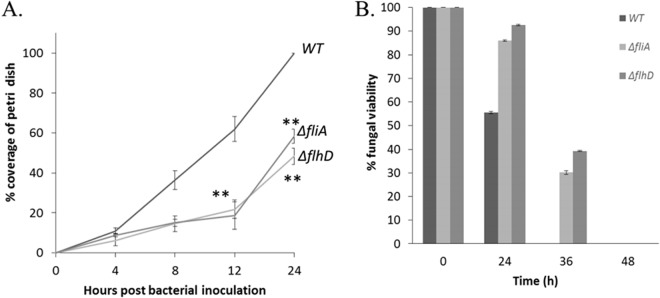
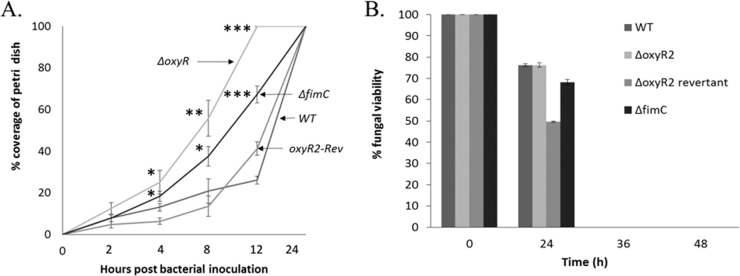
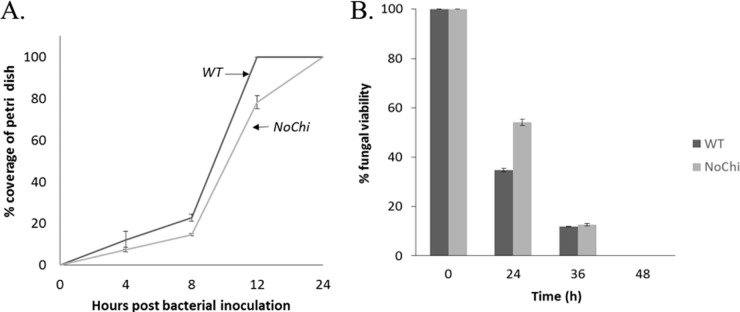
We have found a remarkable capacity for the ubiquitous Gram-negative rod bacterium Serratia marcescens to migrate along and kill the mycelia of zygomycete molds. This migration was restricted to zygomycete molds and several basidiomycete species. No migration was seen on any molds of the phylum Ascomycota. S. marcescens migration did not require fungal viability or surrounding growth medium, as bacteria migrated along aerial hyphae as well.S. marcescens did not exhibit growth tropism toward zygomycete mycelium. Bacterial migration along hyphae proceeded only when the hyphae grew into the bacterial colony. S. marcescens cells initially migrated along the hyphae, forming attached microcolonies that grew and coalesced to generate a biofilm that covered and killed the mycelium. Flagellum-defective strains of S. marcescens were able to migrate along zygomycete hyphae, although they were significantly slower than the wild-type strain and were delayed in fungal killing. Bacterial attachment to the mycelium does not necessitate type 1 fimbrial adhesion, since mutants defective in this adhesin migrated equally well as or faster than the wild-type strain. Killing does not depend on the secretion of S. marcescens chitinases, as mutants in which all three chitinase genes were deleted retained wild-type killing abilities. A better understanding of the mechanisms by which S. marcescens binds to, spreads on, and kills fungal hyphae might serve as an excellent model system for such interactions in general; fungal killing could be employed in agricultural fungal biocontrol.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
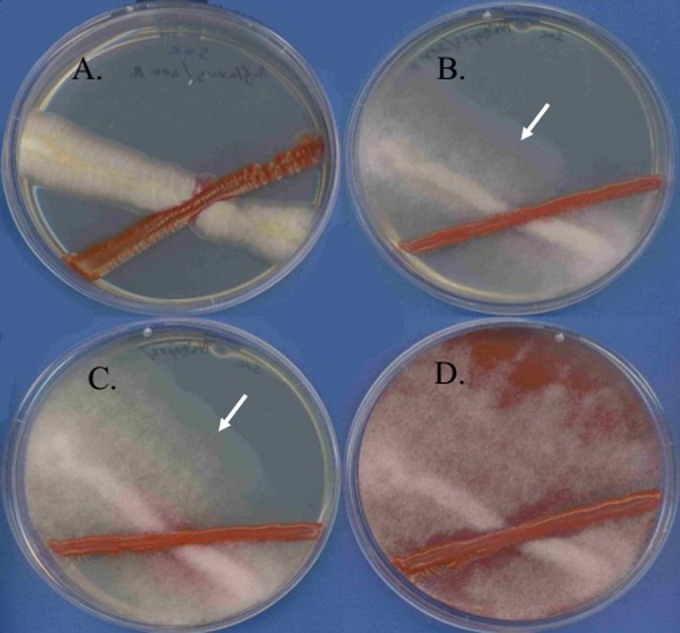
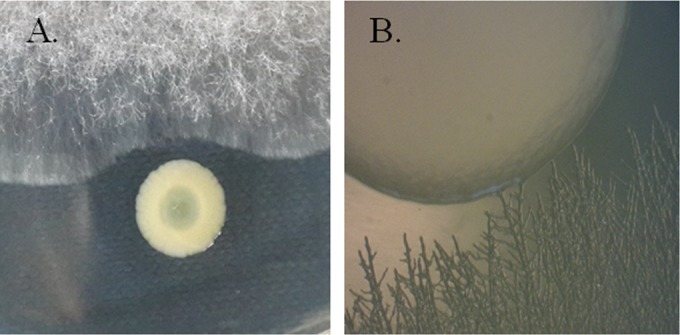
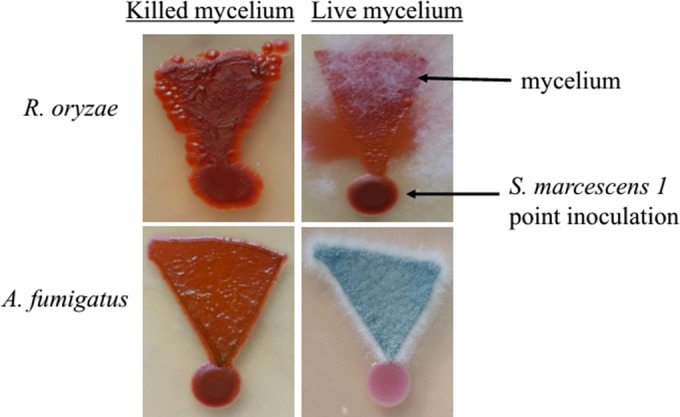

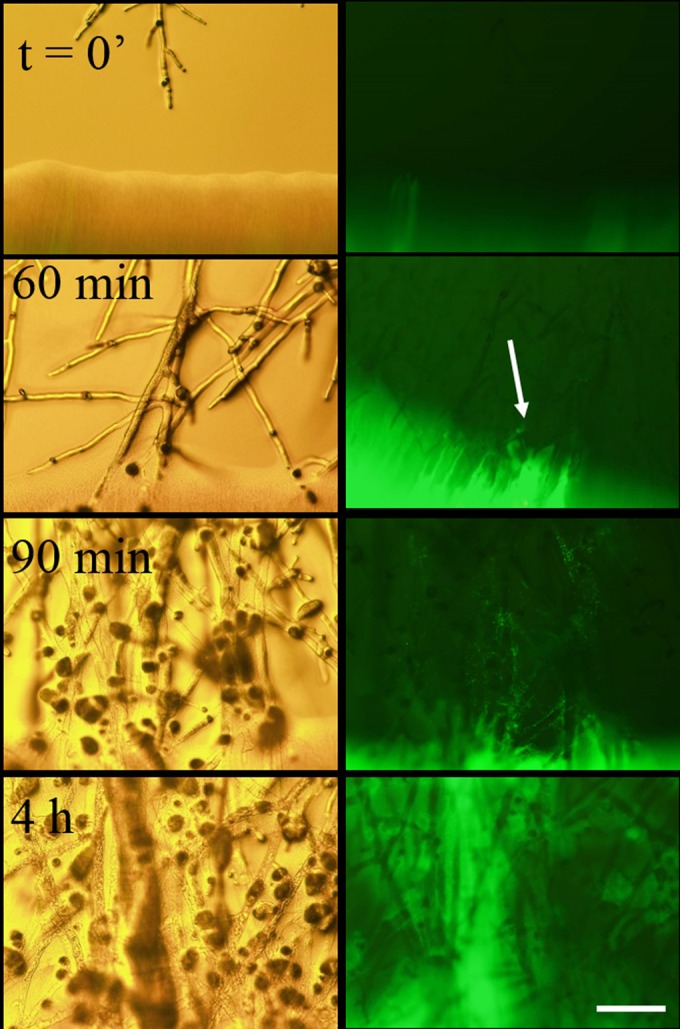
Similar articles
-
Mutualistic interaction of native Serratia marcescens UENF-22GI with Trichoderma longibrachiatum UENF-F476 boosting seedling growth of tomato and papaya.World J Microbiol Biotechnol. 2021 Nov 3;37(12):211. doi: 10.1007/s11274-021-03179-z. World J Microbiol Biotechnol. 2021. PMID: 34729659
-
Mechanism of interaction of an endofungal bacterium Serratia marcescens D1 with its host and non-host fungi.PLoS One. 2020 Apr 22;15(4):e0224051. doi: 10.1371/journal.pone.0224051. eCollection 2020. PLoS One. 2020. PMID: 32320394 Free PMC article.
-
A Serratia marcescens OxyR homolog mediates surface attachment and biofilm formation.J Bacteriol. 2007 Oct;189(20):7262-72. doi: 10.1128/JB.00859-07. Epub 2007 Aug 3. J Bacteriol. 2007. PMID: 17675374 Free PMC article.
-
[Efflux systems in Serratia marcescens].Mikrobiologiia. 2014 Jan-Feb;83(1):3-14. Mikrobiologiia. 2014. PMID: 25423729 Review. Russian.
-
The Mycelium as a Network.Microbiol Spectr. 2017 May;5(3):10.1128/microbiolspec.funk-0033-2017. doi: 10.1128/microbiolspec.FUNK-0033-2017. Microbiol Spectr. 2017. PMID: 28524023 Free PMC article. Review.
Cited by
-
Fungal-Bacterial Interactions in Health and Disease.Pathogens. 2019 May 21;8(2):70. doi: 10.3390/pathogens8020070. Pathogens. 2019. PMID: 31117285 Free PMC article. Review.
-
Antifungal compound from marine Serratia marcescens BKACT and its potential activity against Fusarium sp.Int Microbiol. 2022 Nov;25(4):851-862. doi: 10.1007/s10123-022-00268-3. Epub 2022 Jul 28. Int Microbiol. 2022. PMID: 35900707
-
Genetic Determinants of Antagonistic Interactions and the Response of New Endophytic Strain Serratia quinivorans KP32 to Fungal Phytopathogens.Int J Mol Sci. 2022 Dec 8;23(24):15561. doi: 10.3390/ijms232415561. Int J Mol Sci. 2022. PMID: 36555201 Free PMC article.
-
CpxR-Dependent Thermoregulation of Serratia marcescens PrtA Metalloprotease Expression and Its Contribution to Bacterial Biofilm Formation.J Bacteriol. 2018 Mar 26;200(8):e00006-18. doi: 10.1128/JB.00006-18. Print 2018 Apr 15. J Bacteriol. 2018. PMID: 29378892 Free PMC article.
-
Phenotypic and Proteomic Analysis of the Aspergillus fumigatus ΔPrtT, ΔXprG and ΔXprG/ΔPrtT Protease-Deficient Mutants.Front Microbiol. 2017 Dec 12;8:2490. doi: 10.3389/fmicb.2017.02490. eCollection 2017. Front Microbiol. 2017. PMID: 29312198 Free PMC article.
References
-
- Furuno S, Pazolt K, Rabe C, Neu TR, Harms H, Wick LY. 2010. Fungal mycelia allow chemotactic dispersal of polycyclic aromatic hydrocarbon-degrading bacteria in water-unsaturated systems. Environ Microbiol 12:1391–1398. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
