

Conserved and Diverged Functions of the Calcineurin-Activated Prz1 Transcription Factor in Fission Yeast
- PMID: 26896331
- PMCID: PMC4905549
- DOI: 10.1534/genetics.115.184218
Conserved and Diverged Functions of the Calcineurin-Activated Prz1 Transcription Factor in Fission Yeast
Abstract
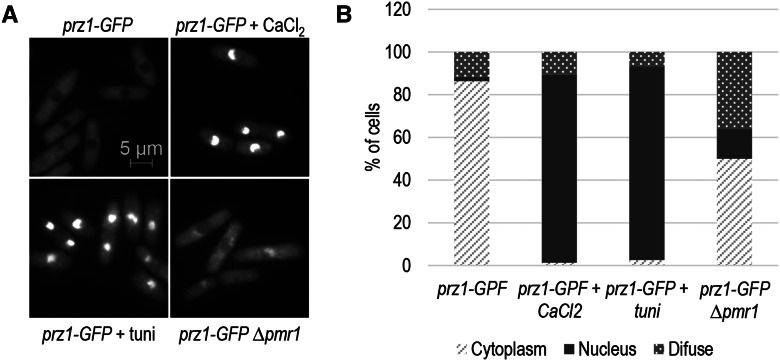
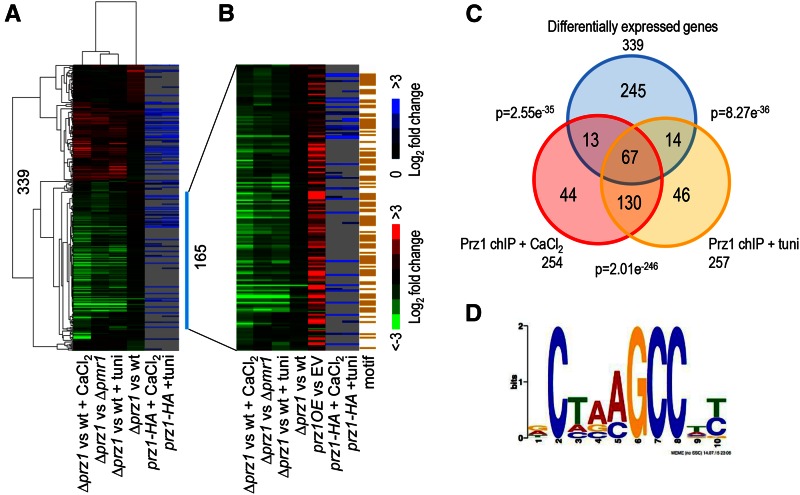
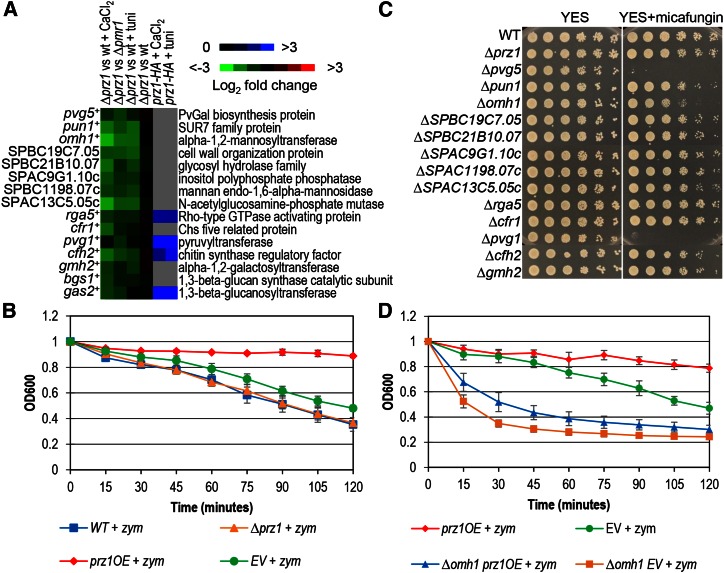
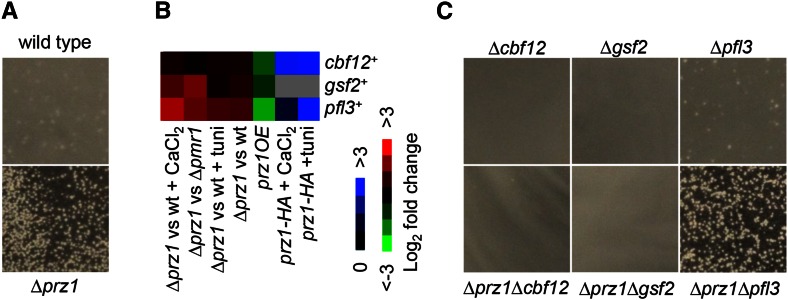
Gene regulation in response to intracellular calcium is mediated by the calcineurin-activated transcription factor Prz1 in the fission yeast Schizosaccharomyces pombe Genome-wide studies of the Crz1 and CrzA fungal orthologs have uncovered numerous target genes involved in conserved and species-specific cellular processes. In contrast, very few target genes of Prz1 have been published. This article identifies an extensive list of genes using transcriptome and ChIP-chip analyses under inducing conditions of Prz1, including CaCl2 and tunicamycin treatment, as well as a ∆pmr1 genetic background. We identified 165 upregulated putative target genes of Prz1 in which the majority contained a calcium-dependent response element in their promoters, similar to that of the Saccharomyces cerevisiae ortholog Crz1 These genes were functionally enriched for Crz1-conserved processes such as cell-wall biosynthesis. Overexpression of prz1(+)increased resistance to the cell-wall degradation enzyme zymolyase, likely from upregulation of theO-mannosyltransferase encoding gene omh1(+) Loss of omh1(+)abrogates this phenotype. We uncovered a novel inhibitory role in flocculation for Prz1. Loss of prz1(+)resulted in constitutive flocculation and upregulation of genes encoding the flocculins Gsf2 and Pfl3, as well as the transcription factor Cbf12. The constitutive flocculation of the ∆prz1 strain was abrogated by the loss of gsf2(+) or cbf12(+) This study reveals that Prz1 functions as a positive and negative transcriptional regulator of genes involved in cell-wall biosynthesis and flocculation, respectively. Moreover, comparison of target genes between Crz1/CrzA and Prz1 indicate some conservation in DNA-binding specificity, but also substantial rewiring of the calcineurin-mediated transcriptional regulatory network.
Keywords: calcium; cell wall; fission yeast; flocculation; transcription factor.
Copyright © 2016 by the Genetics Society of America.
Figures
References
-
- Andreishcheva E. N., Kunkel J. P., Gemmill T. R., Trimble R. B., 2004. Five genes involved in biosynthesis of the pyruvylated Galβ1,3-epitope in Schizosaccharomyces pombe N-linked glycans. J. Biol. Chem. 279: 35644–35655. - PubMed
-
- Araki Y., Wu H., Kitagaki H., Akao T., Takagi H., et al. , 2009. Ethanol stress stimulates the Ca2+-mediated calcineurin/Crz1 pathway in Saccharomyces cerevisiae. J. Biosci. Bioeng. 107: 1–6. - PubMed
-
- Bailey T. L., Elkan C., 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Sec. Int. Conf. Intell. Syst. Mol. Biol. 2: 28–36. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
