

Review
doi: 10.1038/nri.2016.2.
Epub 2016 Feb 22.
Mutations, kataegis and translocations in B cells: understanding AID promiscuous activity
Affiliations
- PMID: 26898111
- PMCID: PMC4871114
- DOI: 10.1038/nri.2016.2
Item in Clipboard
Review
Mutations, kataegis and translocations in B cells: understanding AID promiscuous activity
Nat Rev Immunol.
2016 Mar.
Abstract
As B cells engage in the immune response, they express activation-induced cytidine deaminase (AID) to initiate the hypermutation and recombination of immunoglobulin genes, which are crucial processes for the efficient recognition and disposal of pathogens. However, AID must be tightly controlled in B cells to minimize off-target mutations, which can drive chromosomal translocations and the development of B cell malignancies, such as lymphomas. Recent genomic and biochemical analyses have begun to unravel the mechanisms of how AID-mediated deamination is targeted outside immunoglobulin genes. Here, we discuss the transcriptional and topological features that are emerging as key drivers of AID promiscuous activity.
Figures
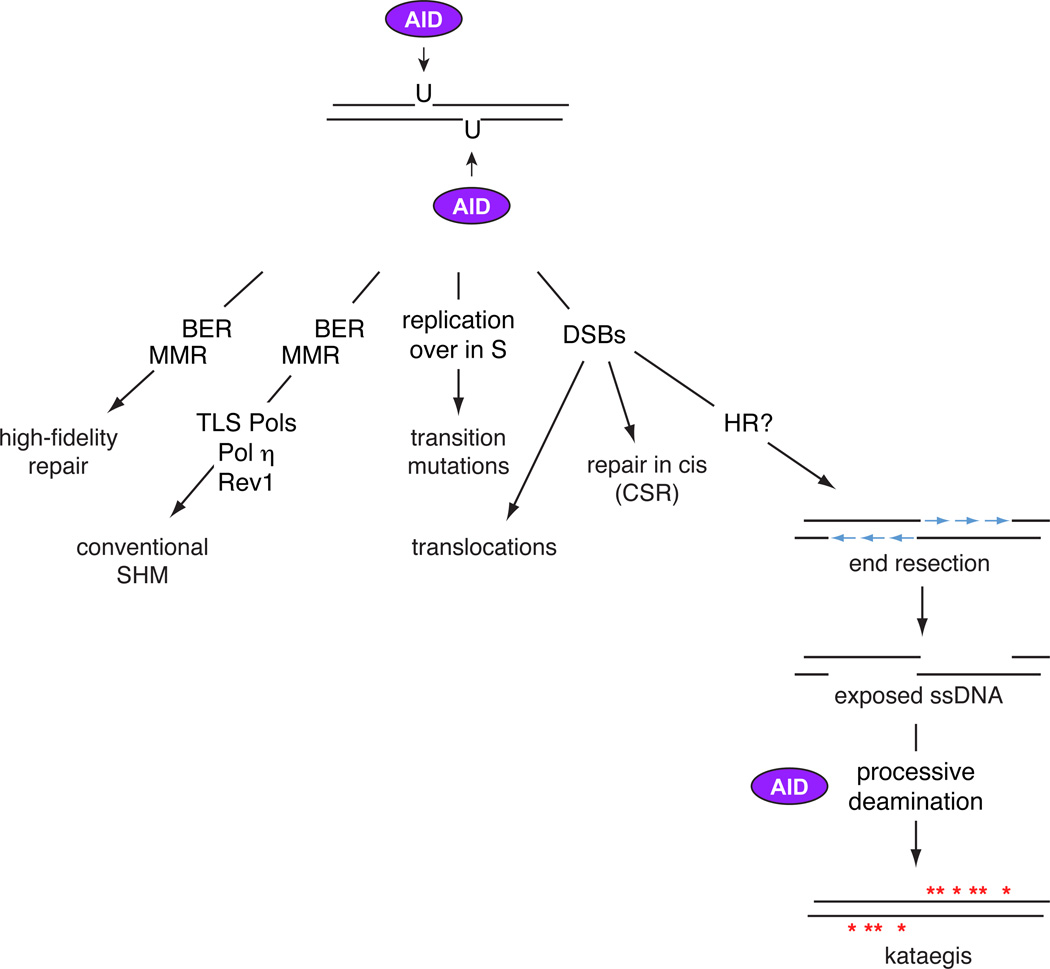
The deamination of genomic DNA by AID can be processed in at least 5 distinct modes. a) High-fidelity repair by BER and MMR factors can revert deoxyuridines to deoxycitidines. b) The BER and MMR pathways, in combination with translesion polymerases such as Rev1 and Polη, carry out conventional SHM. c) DNA replication over deoxyuridines in the S phase of the cell cycle leads to transition mutations. d) The formation of staggered DSBs are either repaired in cis by NHEJ leading to CSR, or in trans leading to chromosomal translocations. e) If DNA breaks are unrepaired as the cell moves to the S or G2M stages of the cell cycle, attempts to initiate homologous recombination promotes the resection of DNA ends. The resulting ssDNA are potential targets to AID, which by processive deamination may generate kataegis.
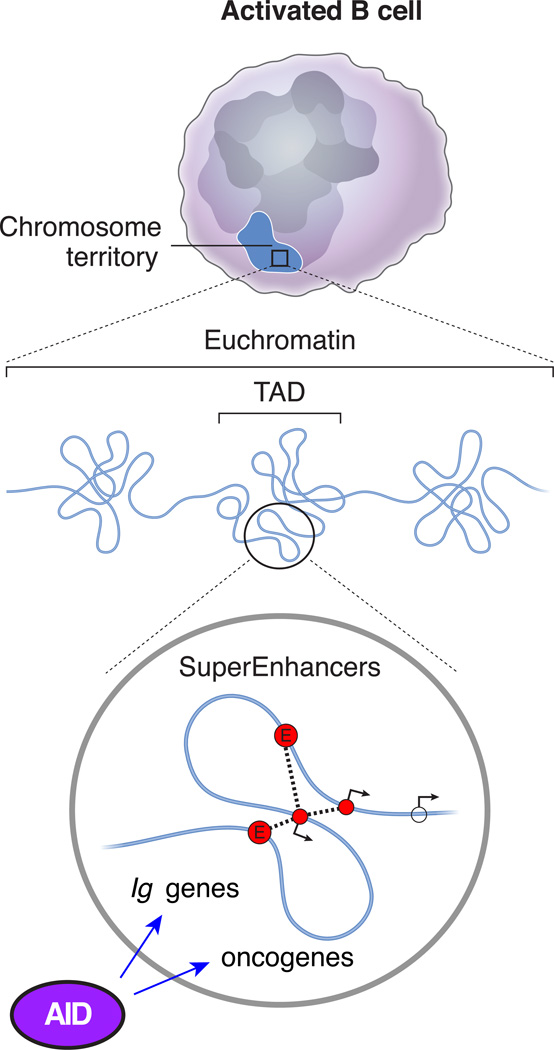
Topological, transcriptional, and epigenetic features that render super-enhancer domains ideal targets for AID activity, both at immunoglobulin loci and at selected oncogenes in B cells, as well as in somatic cells when AID is ectopically expressed.
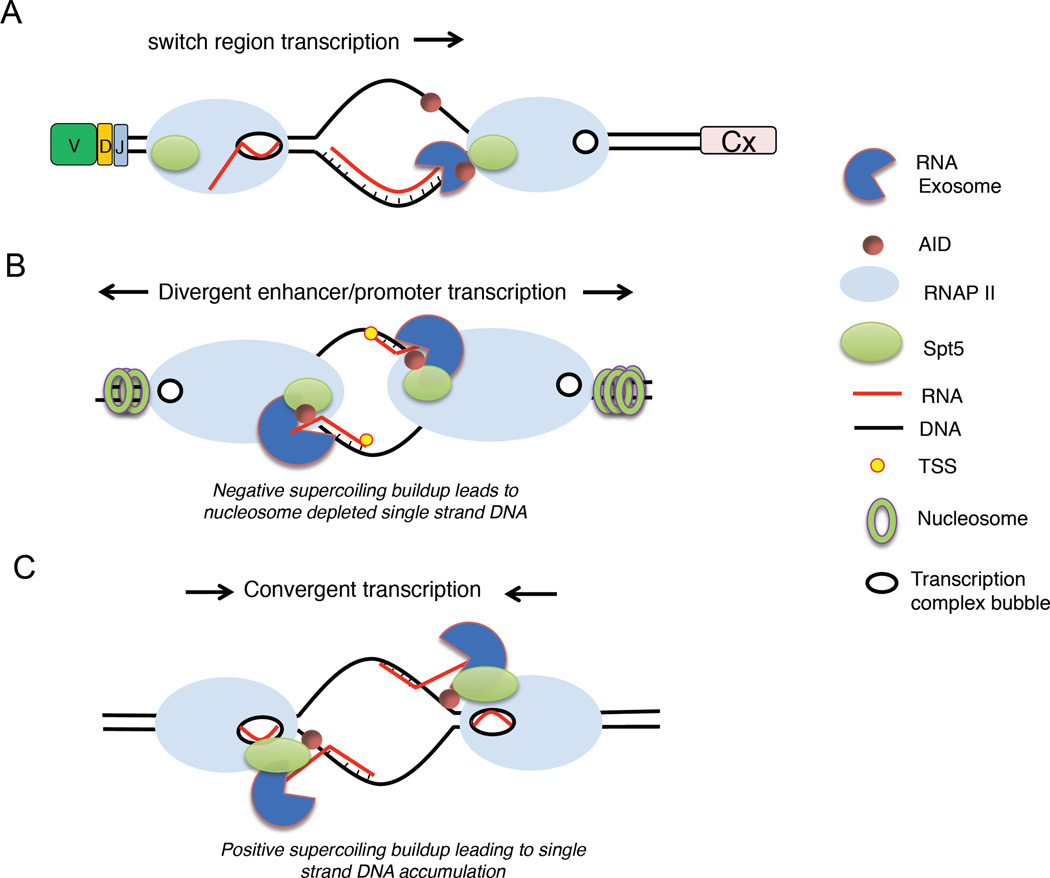
(A) Transcription of S regions creates R-loops that cause stalling of RNA pol II, early transcription termination and RNA exosome recruitment. (B) Divergent transcription at enhancers and promoters create nucleosome-free DNA and ssDNA structures where cognate RNAs may associate and become substrates for exosome degradation. These activities are proposed to increase accessibility to AID. (C) Convergent transcription mediated by pol II may lead to the formation of RNA exosome substrates by the buildup of positive DNA supercoiling. SPT5, transcription elongation factor SPT5; TSS, transcription start site.
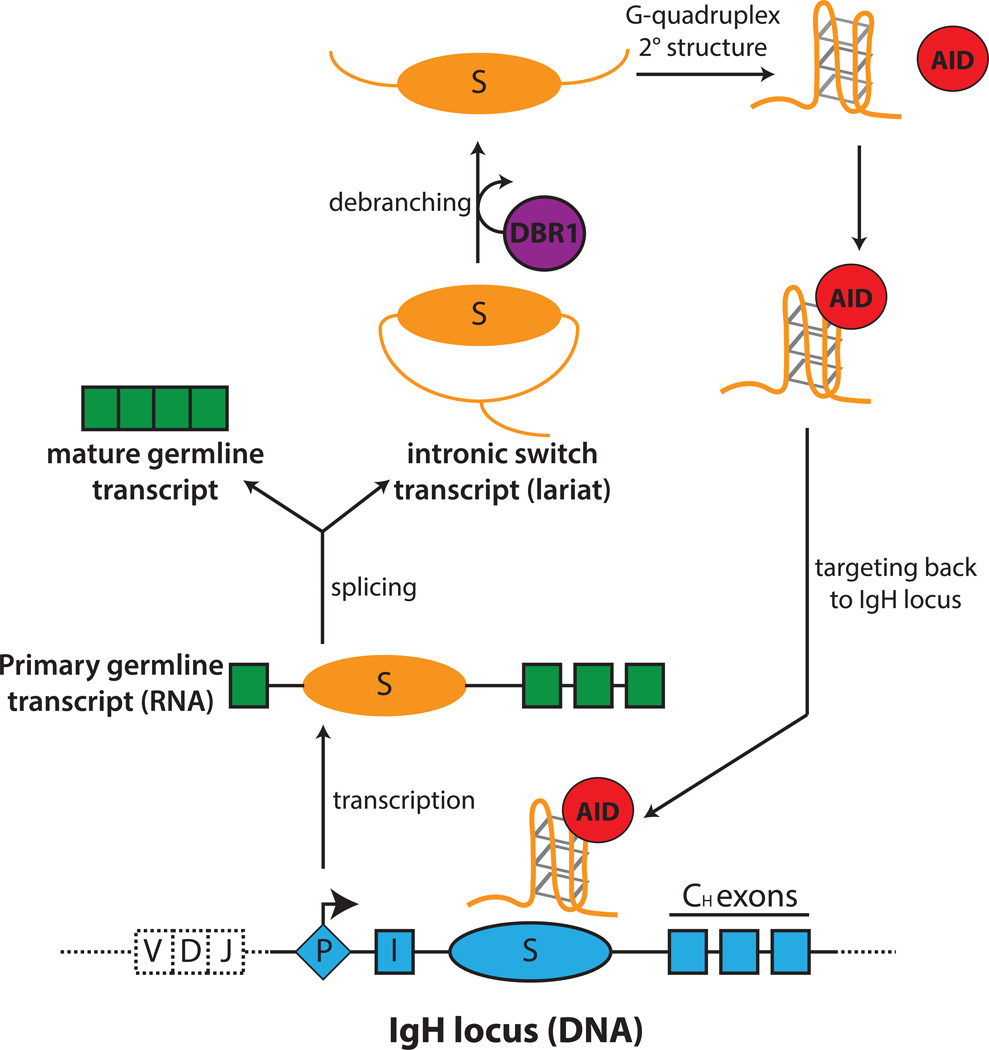
When B cells are stimulated to undergo CSR, transcription occurs at each of the recombining switch (S) regions to produce primary switch transcripts. Primary transcripts are spliced to generate a mature germline transcripts and intronic switch region transcripts (lariat intermediate). Debranching enzyme 1 (DBR1) catalyzes the release of the lariat from the spliceosome and debranches the switch transcript into its linear form. The linear switch transcript, free of exonic sequences, can function as a guide RNA by forming a G-quadruplex structure, which allows its association with AID. AID, bound to the guide RNA, is targeted specifically by sequence information provided by the guide RNAs to the complementary S region DNA.
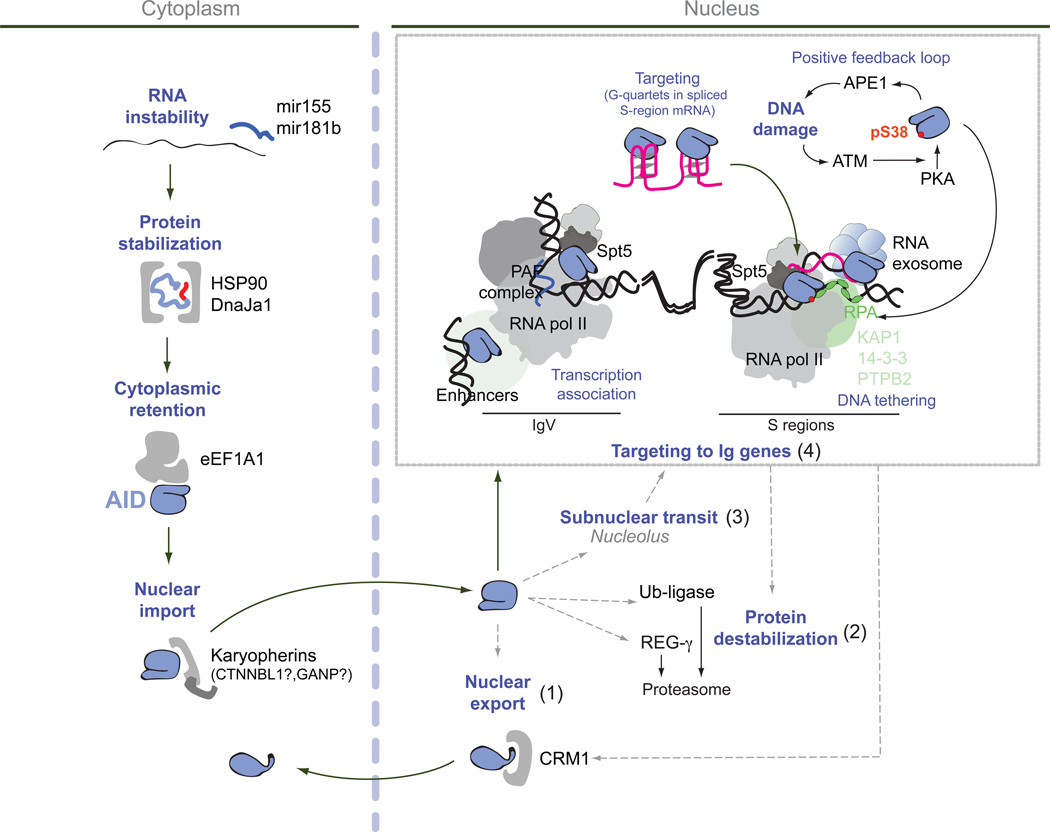
AID transits through HSP90- and eEF1A–containing complexes in the cytoplasm that control its functional maturation before being imported into the nucleus. Whether nuclear import of AID is regulated or stochastic is unknown. In the nucleus AID has multiple possible destinations. It can be: one, exported back to the cytoplasm, two, destabilized by proteasomal degradation, three, associate with the nucleolus and four, be recruited to the chromatin of immunoglobulin variable (IgV) and/or S regions as well as to off-target genes. It remains to be determined to what extent these various destinations are sequential while AID travels to the Ig loci, or compete for AID or synergize to minimize off-targeting. Targeting of AID to the IgV or S-regions requires strong transcription by RNA polymerase II associated with Spt5 and the PAF complex in the context of a specific chromatin microenvironment that includes abundant topological associations of multiple enhancers. Transcription can lead to DNA supercoiling and expose ssDNA that is stabilized by RPA. The repetitiveness of S-regions facilitates the formation of R-loops, exposing the untranscribed DNA strand. The exosome helps recruit AID to the template strand of R loops by degrading R-loop associated transcripts. AID is also recruited by genomic regions displaying extensive convergent or divergent transcription. A number of chromatin associating factors might help tether AID to chromatin, at least at S regions. These include KAP1, 14-3-3 and PTBP2. Once recruited, AID initiates a series of events leading to the formation of DNA breaks (after excision of uracil by UNG and the DNA nicking activity of the endonuclease APE1), which activates the DNA damage response kinase ATM. ATM promotes PKA-mediated phosphorylation of AID at serine 38. This phosphorylation increases AID activity and allows AID association to RPA. In addition, AID binds to G-quartet structures formed in the debranched intron of the S-region sterile transcript, which presumable pairs back with the DNA and helps targeting AID for CSR. Black arrows indicate mechanisms thought to act in chronological order and/or that are directly linked. Dashed grey arrows indicate possible but not yet empirically demonstrated connections.
References
-
- Pieper K, Grimbacher B, Eibel H. B-cell biology and development. J Allergy Clin Immunol. 2013;131:959–971. - PubMed
-
- Di Noia JM, Neuberger MS. Molecular mechanisms of antibody somatic hypermutation. Annu Rev Biochem. 2007;76:1–22. - PubMed
-
- Revy P, et al. Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the Hyper-IgM syndrome (HIGM2)[see comments] Cell. 2000;102:565–575. - PubMed
-
- Muramatsu M, et al. Class switch recombination hypermutation require activation-induced cytidine deaminase (AID), a potential RNAediting enzyme [see comments] Cell. 2000;102:553–563. - PubMed
-
- Shen HM, Peters A, Baron B, Zhu X, Storb U. Mutation of BCL-6 gene in normal B cells by the process of somatic hypermutation of Ig genes. Science. 1998;280:1750–1752. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
