

Chromosome Cohesion Established by Rec8-Cohesin in Fetal Oocytes Is Maintained without Detectable Turnover in Oocytes Arrested for Months in Mice
- PMID: 26898469
- PMCID: PMC4791431
- DOI: 10.1016/j.cub.2015.12.073
Chromosome Cohesion Established by Rec8-Cohesin in Fetal Oocytes Is Maintained without Detectable Turnover in Oocytes Arrested for Months in Mice
Abstract
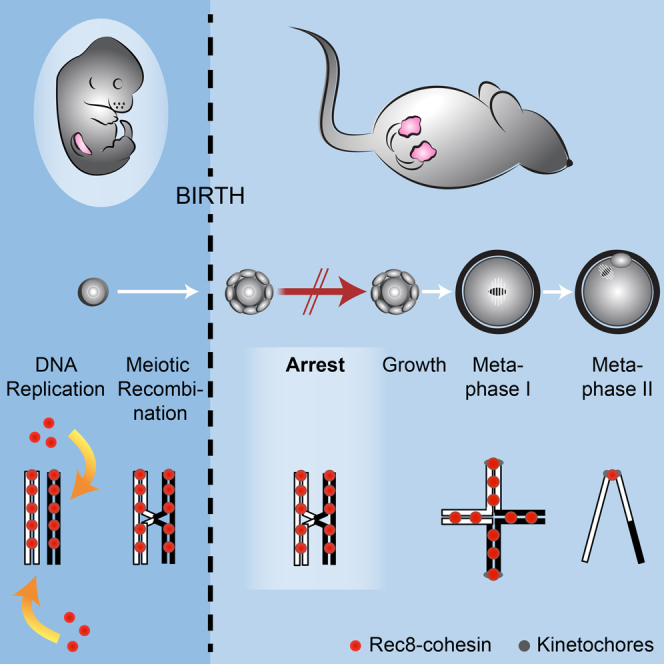
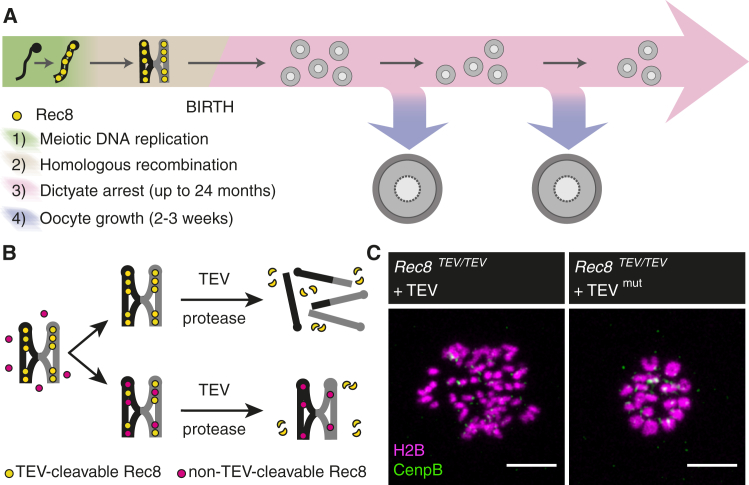
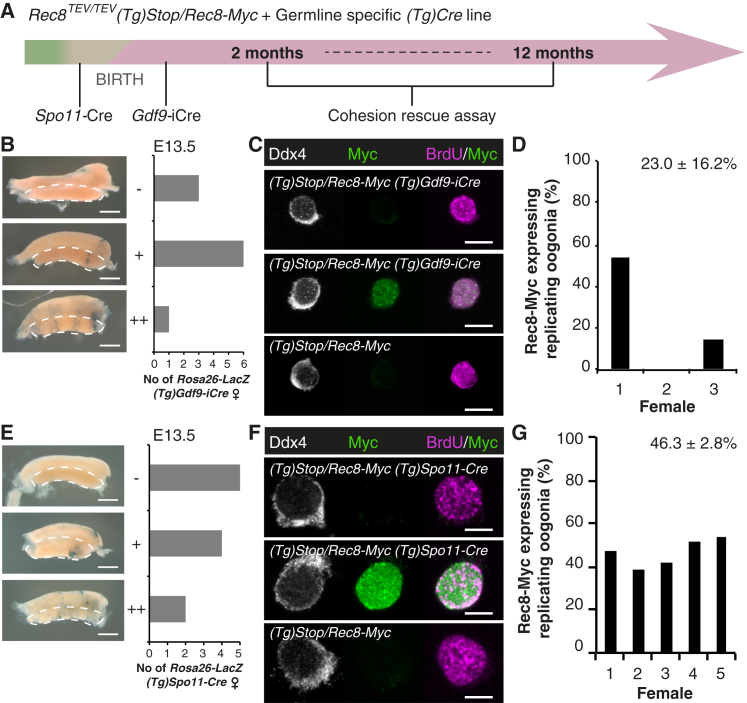
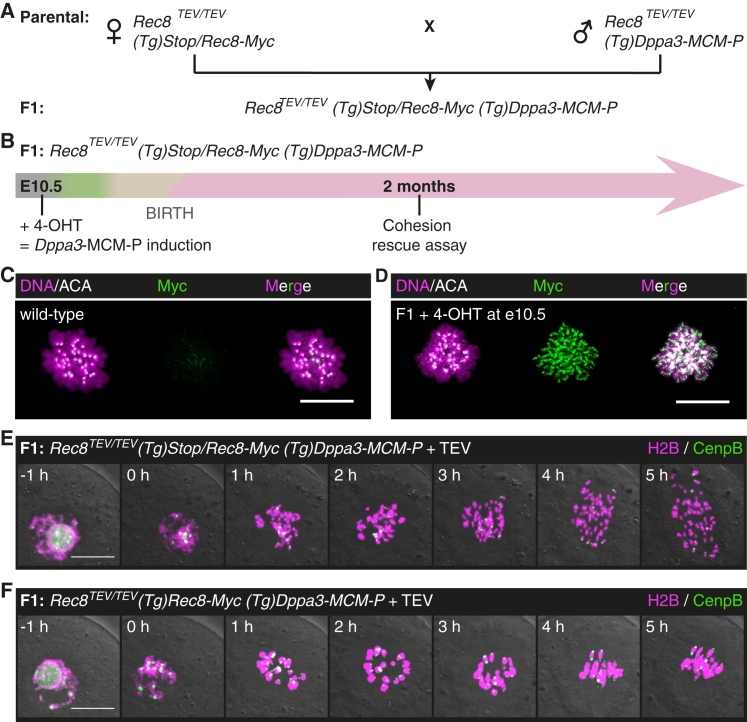
Sister chromatid cohesion mediated by the cohesin complex is essential for chromosome segregation in mitosis and meiosis [1]. Rec8-containing cohesin, bound to Smc3/Smc1α or Smc3/Smc1β, maintains bivalent cohesion in mammalian meiosis [2-6]. In females, meiotic DNA replication and recombination occur in fetal oocytes. After birth, oocytes arrest at the prolonged dictyate stage until recruited to grow into mature oocytes that divide at ovulation. How cohesion is maintained in arrested oocytes remains a pivotal question relevant to maternal age-related aneuploidy. Hypothetically, cohesin turnover regenerates cohesion in oocytes. Evidence for post-replicative cohesion establishment mechanism exists, in yeast and invertebrates [7, 8]. In mouse fetal oocytes, cohesin loading factor Nipbl/Scc2 localizes to chromosome axes during recombination [9, 10]. Alternatively, cohesion is maintained without turnover. Consistent with this, cohesion maintenance does not require Smc1β transcription, but unlike Rec8, Smc1β is not required for establishing bivalent cohesion [11, 12]. Rec8 maintains cohesion without turnover during weeks of oocyte growth [3]. Whether the same applies to months or decades of arrest is unknown. Here, we test whether Rec8 activated in arrested mouse oocytes builds cohesion revealed by TEV cleavage and live-cell imaging. Rec8 establishes cohesion when activated during DNA replication in fetal oocytes using tamoxifen-inducible Cre. In contrast, no new cohesion is detected when Rec8 is activated in arrested oocytes by tamoxifen despite cohesin synthesis. We conclude that cohesion established in fetal oocytes is maintained for months without detectable turnover in dictyate-arrested oocytes. This implies that women's fertility depends on the longevity of cohesin proteins that established cohesion in utero.
Keywords: cohesin; meiosis; oocytes.
Copyright © 2016 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
Comment in
-
Oogenesis: Ageing Oocyte Chromosomes Rely on Amazing Protein Stability.Curr Biol. 2016 Apr 25;26(8):R329-31. doi: 10.1016/j.cub.2016.02.059. Curr Biol. 2016. PMID: 27115691
References
-
- Nasmyth K., Haering C.H. Cohesin: its roles and mechanisms. Annu. Rev. Genet. 2009;43:525–558. - PubMed
-
- Watanabe Y., Nurse P. Cohesin Rec8 is required for reductional chromosome segregation at meiosis. Nature. 1999;400:461–464. - PubMed
-
- Lee J., Iwai T., Yokota T., Yamashita M. Temporally and spatially selective loss of Rec8 protein from meiotic chromosomes during mammalian meiosis. J. Cell Sci. 2003;116:2781–2790. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
