

Genome analysis of three Pneumocystis species reveals adaptation mechanisms to life exclusively in mammalian hosts
- PMID: 26899007
- PMCID: PMC4764891
- DOI: 10.1038/ncomms10740
Genome analysis of three Pneumocystis species reveals adaptation mechanisms to life exclusively in mammalian hosts
Abstract
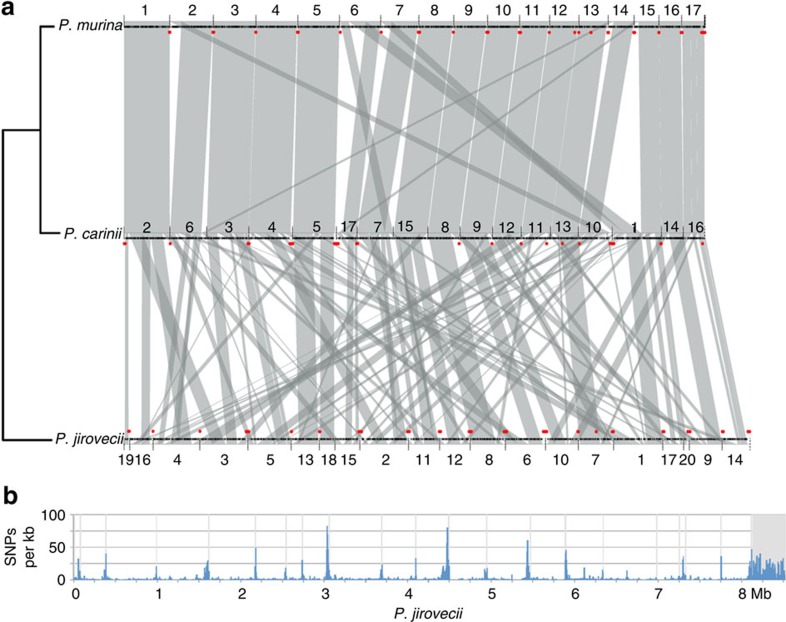
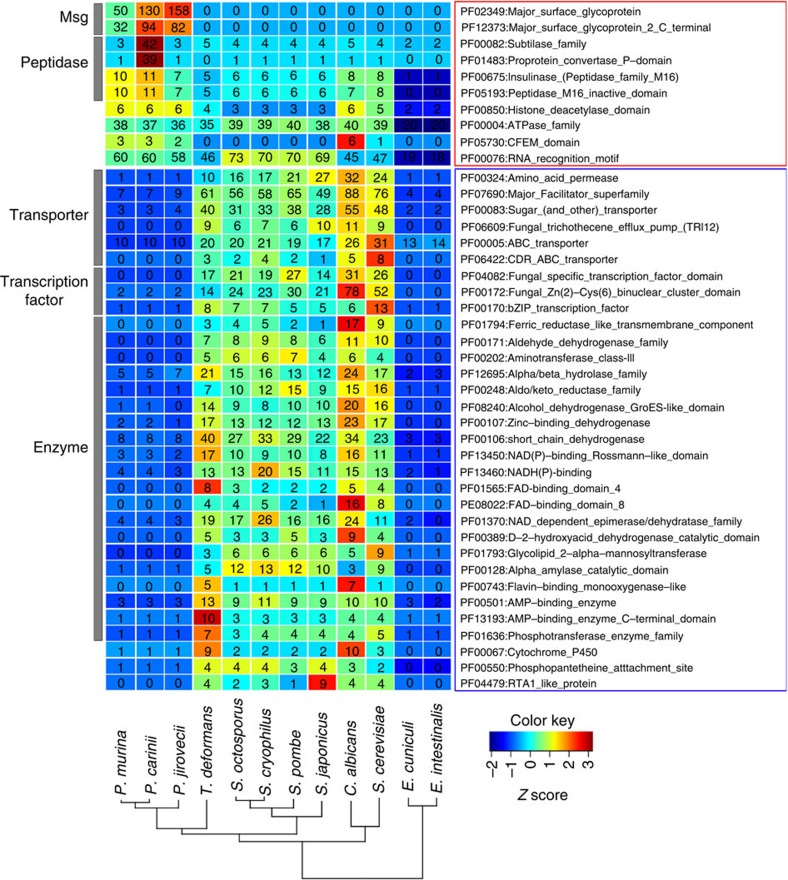
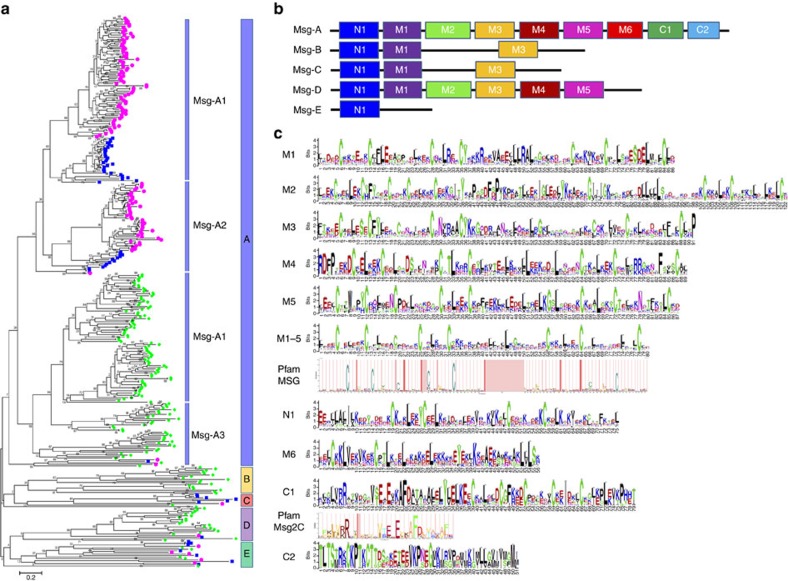
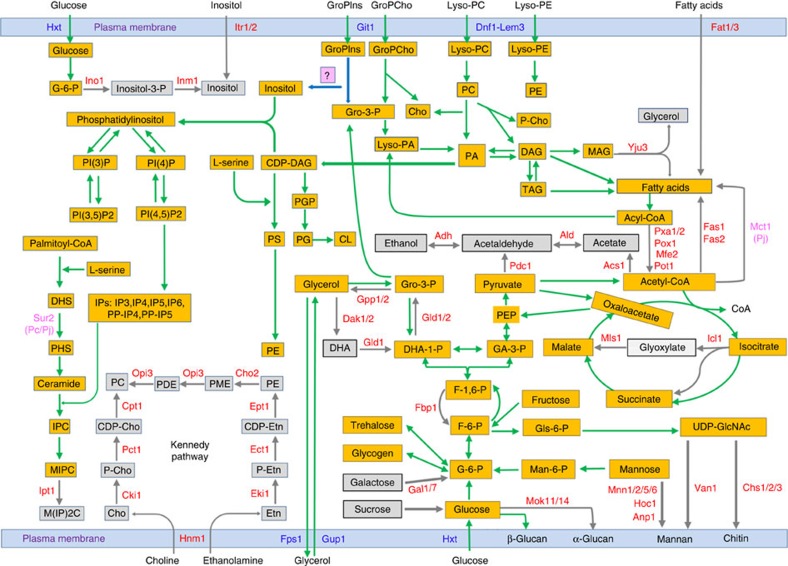
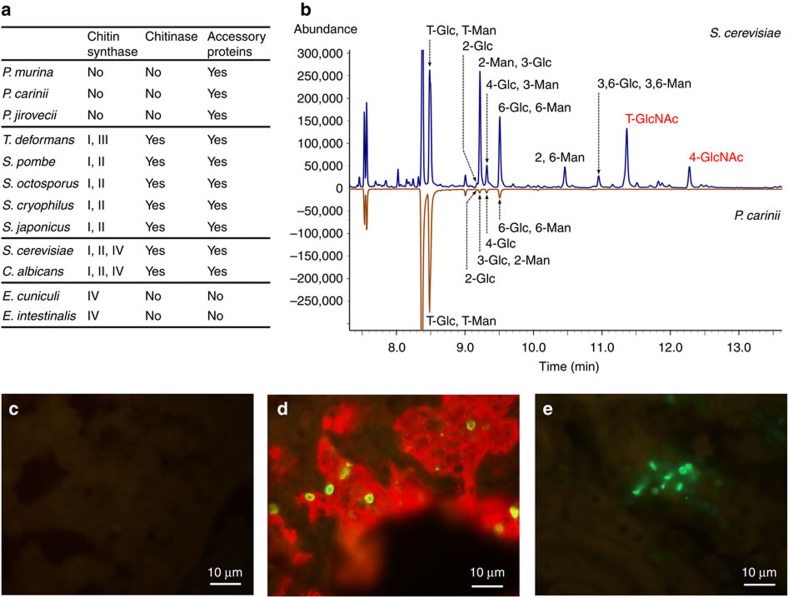
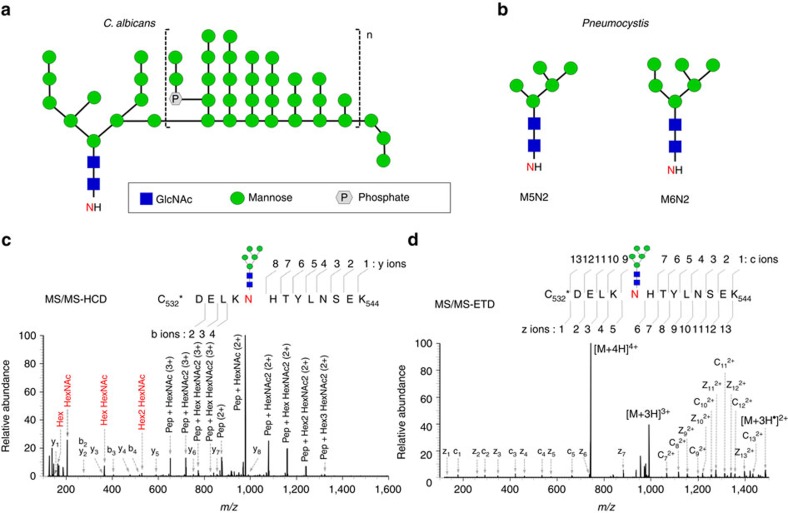
Pneumocystis jirovecii is a major cause of life-threatening pneumonia in immunosuppressed patients including transplant recipients and those with HIV/AIDS, yet surprisingly little is known about the biology of this fungal pathogen. Here we report near complete genome assemblies for three Pneumocystis species that infect humans, rats and mice. Pneumocystis genomes are highly compact relative to other fungi, with substantial reductions of ribosomal RNA genes, transporters, transcription factors and many metabolic pathways, but contain expansions of surface proteins, especially a unique and complex surface glycoprotein superfamily, as well as proteases and RNA processing proteins. Unexpectedly, the key fungal cell wall components chitin and outer chain N-mannans are absent, based on genome content and experimental validation. Our findings suggest that Pneumocystis has developed unique mechanisms of adaptation to life exclusively in mammalian hosts, including dependence on the lungs for gas and nutrients and highly efficient strategies to escape both host innate and acquired immune defenses.
Figures

References
-
- Thomas C. F. Jr & Limper A. H. Current insights into the biology and pathogenesis of Pneumocystis pneumonia. Nat. Rev. Microbiol. 5, 298–308 (2007). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
