

Identification of Novel Candidate Genes for Early-Onset Colorectal Cancer Susceptibility
- PMID: 26901136
- PMCID: PMC4764646
- DOI: 10.1371/journal.pgen.1005880
Identification of Novel Candidate Genes for Early-Onset Colorectal Cancer Susceptibility
Abstract
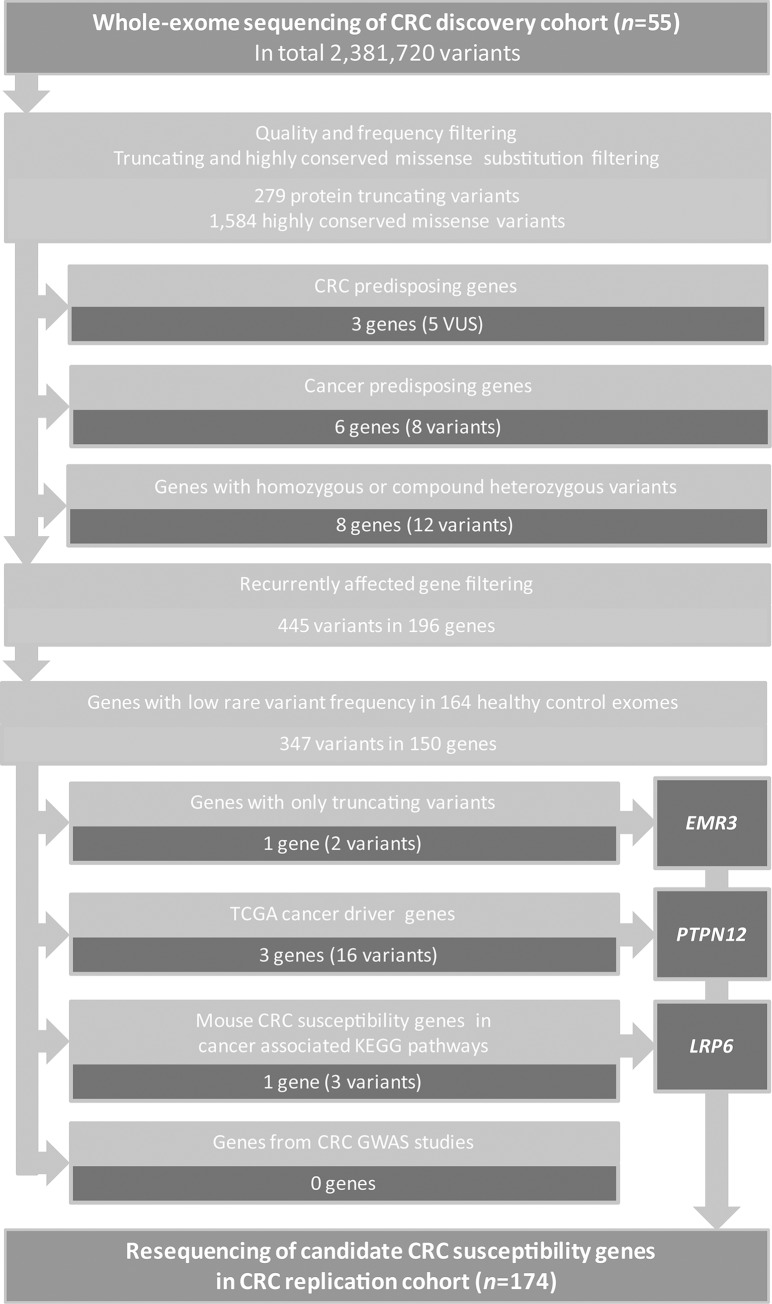
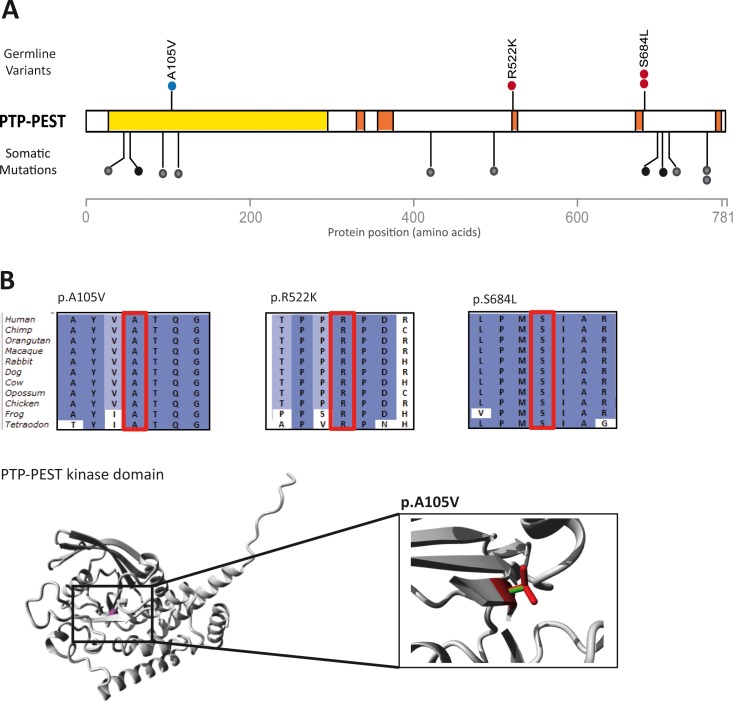
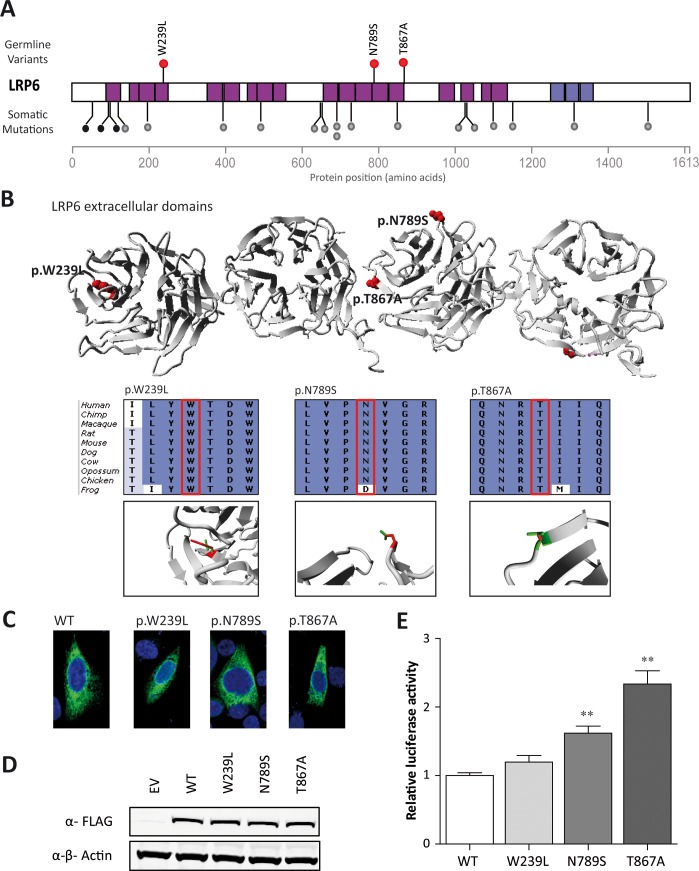
Approximately 25-30% of colorectal cancer (CRC) cases are expected to result from a genetic predisposition, but in only 5-10% of these cases highly penetrant germline mutations are found. The remaining CRC heritability is still unexplained, and may be caused by a hitherto-undefined set of rare variants with a moderately penetrant risk. Here we aimed to identify novel risk factors for early-onset CRC using whole-exome sequencing, which was performed on a cohort of CRC individuals (n = 55) with a disease onset before 45 years of age. We searched for genes that were recurrently affected by rare variants (minor allele frequency ≤ 0.001) with potentially damaging effects and, subsequently, re-sequenced the candidate genes in a replication cohort of 174 early-onset or familial CRC individuals. Two functionally relevant genes with low frequency variants with potentially damaging effects, PTPN12 and LRP6, were found in at least three individuals. The protein tyrosine phosphatase PTP-PEST, encoded by PTPN12, is a regulator of cell motility and LRP6 is a component of the WNT-FZD-LRP5-LRP6 complex that triggers WNT signaling. All variants in LRP6 were identified in individuals with an extremely early-onset of the disease (≤30 years of age), and two of the three variants showed increased WNT signaling activity in vitro. In conclusion, we present PTPN12 and LRP6 as novel candidates contributing to the heterogeneous susceptibility to CRC.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
