

The successful induction of T-cell and antibody responses by a recombinant measles virus-vectored tetravalent dengue vaccine provides partial protection against dengue-2 infection
- PMID: 26901482
- PMCID: PMC4964840
- DOI: 10.1080/21645515.2016.1143576
The successful induction of T-cell and antibody responses by a recombinant measles virus-vectored tetravalent dengue vaccine provides partial protection against dengue-2 infection
Abstract
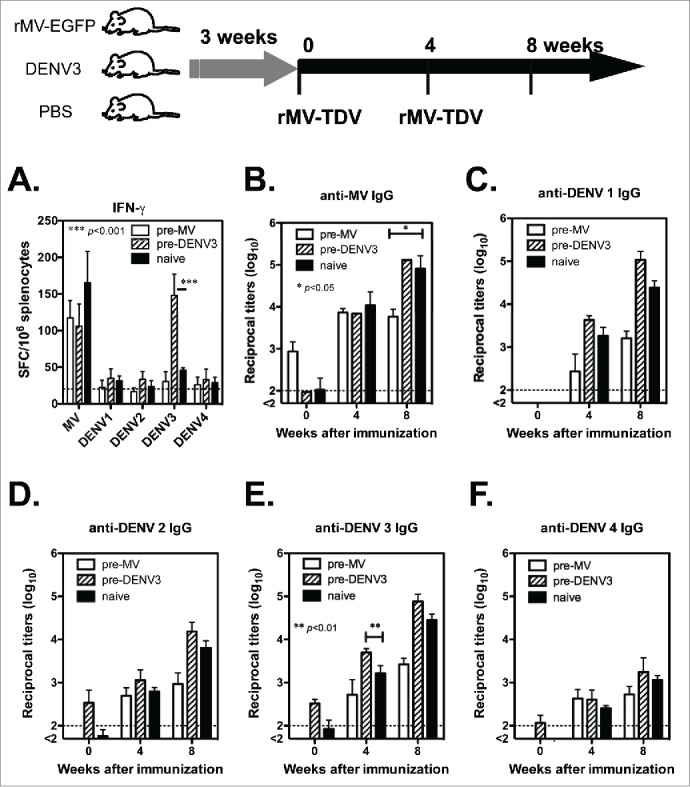
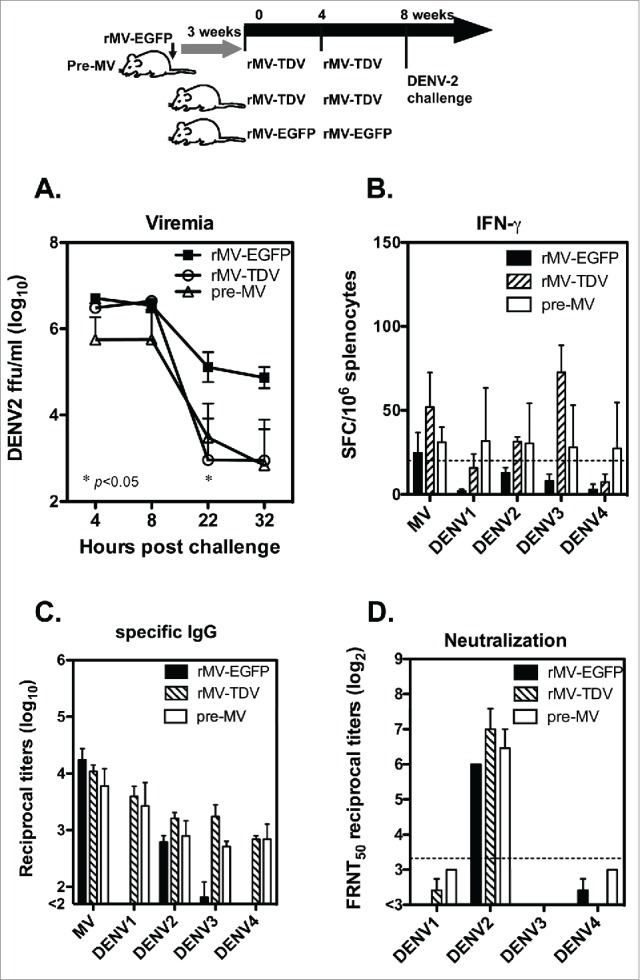
Dengue has a major impact on global public health, and the use of dengue vaccine is very limited. In this study, we evaluated the immunogenicity and protective efficacy of a dengue vaccine made from a recombinant measles virus (MV) that expresses envelope protein domain III (ED3) of dengue-1 to 4. Following immunization with the MV-vectored dengue vaccine, mice developed specific interferon-gamma and antibody responses against dengue virus and MV. Neutralizing antibodies against MV and dengue viruses were also induced, and protective levels of FRNT50 ≥ 10 to 4 serotypes of dengue viruses were detected in the MV-vectored dengue vaccine-immunized mice. In addition, specific interferon-gamma and antibody responses to dengue viruses were still induced by the MV-vectored dengue vaccine in mice that were pre-infected with MV. This finding suggests that the pre-existing immunity to MV did not block the initiation of immune responses. By contrast, mice that were pre-infected with dengue-3 exhibited no effect in terms of their antibody responses to MV and dengue viruses, but a dominant dengue-3-specific T-cell response was observed. After injection with dengue-2, a detectable but significantly lower viremia and a higher titer of anti-dengue-2 neutralizing antibodies were observed in MV-vectored dengue vaccine-immunized mice versus the vector control, suggesting that an anamnestic antibody response that provided partial protection against dengue-2 was elicited. Our results with regard to T-cell responses and the effect of pre-immunity to MV or dengue viruses provide clues for the future applications of an MV-vectored dengue vaccine.
Figures
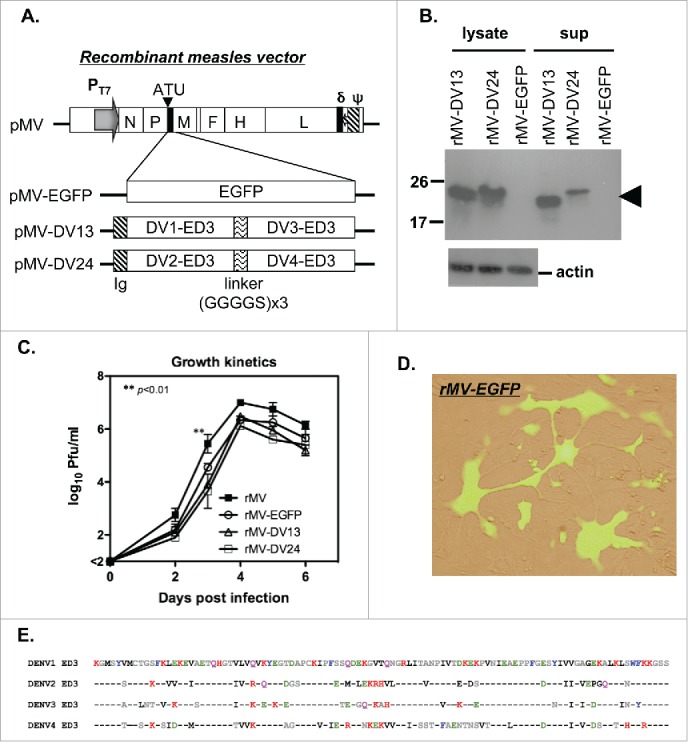
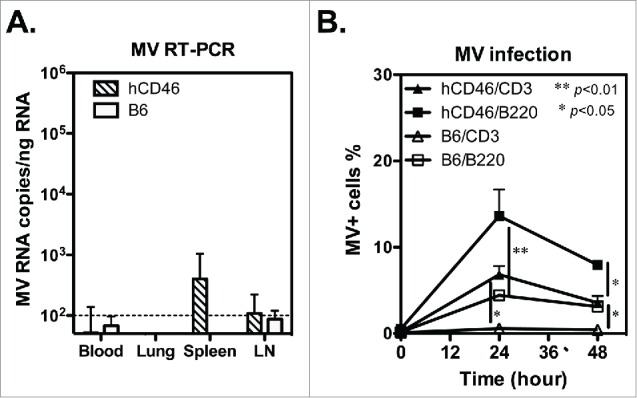
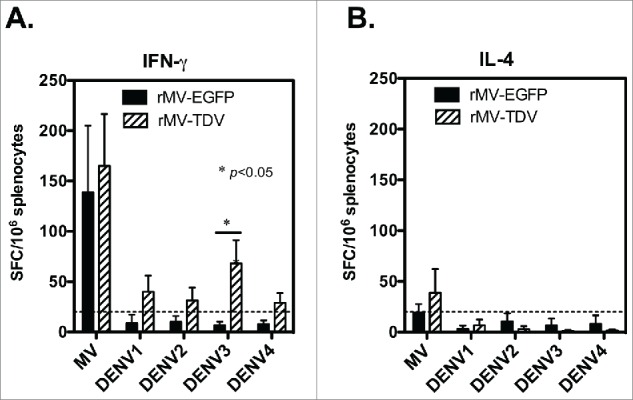
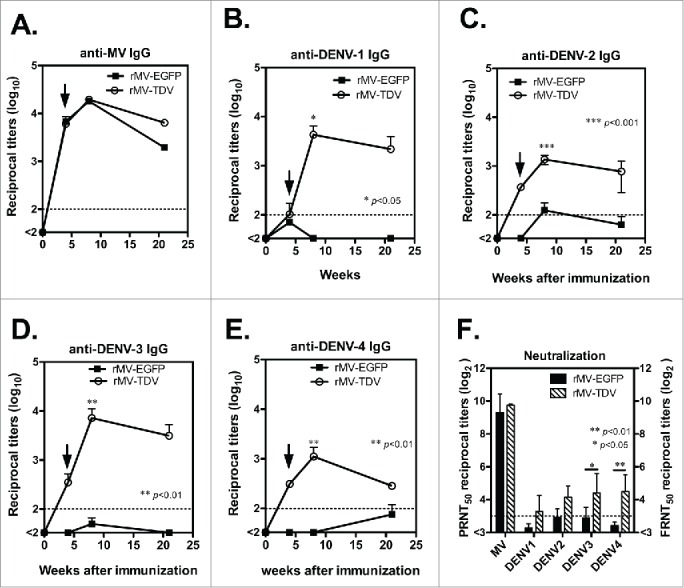
References
-
- Thomas SJ, Endy TP. Vaccines for the prevention of dengue: development update. Hum Vaccin 2011; 7:674-84; PMID:21508679; http://dx.doi.org/ 10.4161/hv.7.6.14985 - DOI - PubMed
-
- Ramasamy R, Surendran SN. Global climate change and its potential impact on disease transmission by salinity-tolerant mosquito vectors in coastal zones. Front Physiol 2012; 3:198; PMID:22723781; http://dx.doi.org/ 10.3389/fphys.2012.00198 - DOI - PMC - PubMed
-
- Sabin AB. Research on dengue during World War II. Am J Trop Med Hyg 1952; 1:30-50; PMID:14903434 - PubMed
-
- Innis BL. Antibody responses to dengue virus infection. New York: CAB Int., 1997.
-
- Halstead SB. Pathogenesis of dengue: challenges to molecular biology. Science 1988; 239:476-81; PMID:3277268; http://dx.doi.org/ 10.1126/science.3277268 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources