

doi: 10.1038/nmeth.3771.
Epub 2016 Feb 22.
High-resolution mass spectrometry of small molecules bound to membrane proteins
Affiliations
- PMID: 26901650
- PMCID: PMC4856209
- DOI: 10.1038/nmeth.3771
Item in Clipboard
High-resolution mass spectrometry of small molecules bound to membrane proteins
Nat Methods.
2016 Apr.
Abstract
Small molecules are known to stabilize membrane proteins and to modulate their function and oligomeric state, but such interactions are often hard to precisely define. Here we develop and apply a high-resolution, Orbitrap mass spectrometry-based method for analyzing intact membrane protein-ligand complexes. Using this platform, we resolve the complexity of multiple binding events, quantify small molecule binding and reveal selectivity for endogenous lipids that differ only in acyl chain length.
Figures
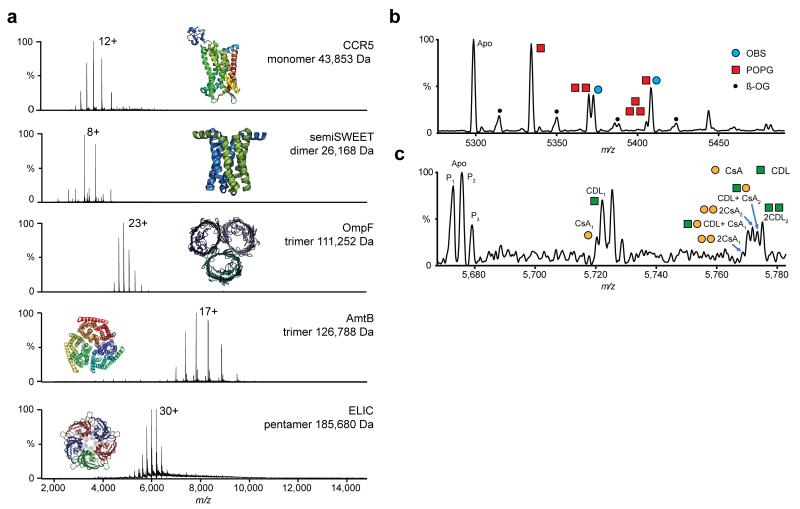
Non-denaturing Orbitrap mass spectra of purified membrane protein complexes of differing size and topology, bound to multiple lipids, peptides and drugs. (a) High resolution non-denaturing Orbitrap mass spectra of the monomeric GPCR chemokine receptor CCR5, dimeric glycan transporter semiSWEET, trimeric outer membrane porin OmpF, trimeric ammonia transporter AmtB and the pentameric ligand gated ion channel ELIC. Masses and associated errors can be found in Supplementary Table 1. Protein structures have been generated from PDB files 4QND, 4MBS, 2NPE, 2OMF and 3RQU. (b,c) Concomitant small molecule binding. (b) Binding of OBS (blue circle) to OmpF in the presence of POPG lipid (red square). Expansion of the 22+ charge state. The OBS peak can be clearly separated from the 2POPG peak. (c) Binding of CsA drug (yellow circle) to P-gp in the presence of CDL 14:0 (green square). Zoom of two ligand binding events on the 25+ charge state. P-gp is present in three proteoforms (denoted by subscript). Both CsA and CDL binding can be clearly resolved. For the peak corresponding to two ligands bound, fine structure indicates the binding of different numbers of CsA and CDL to distinct proteoforms of P-gp. (For detailed assignment see Supplementary Table 2 and for individual ligand binding Supplementary Figure 7).
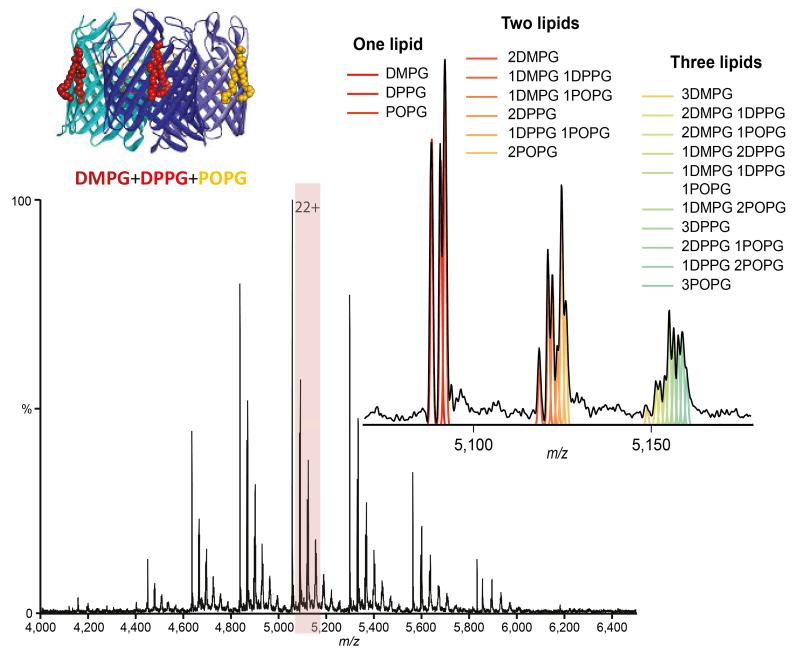
Mass spectrum of trimeric OmpF bound to an equimolar ratio of DMPG, DPPG and POPG lipids (cartoon inset) and zoom of the 22+ charge state with peaks showing up to three lipids bound. Theoretical distributions corresponding to different combinations of lipids are shown by coloured lines and correlate with the spectrum.
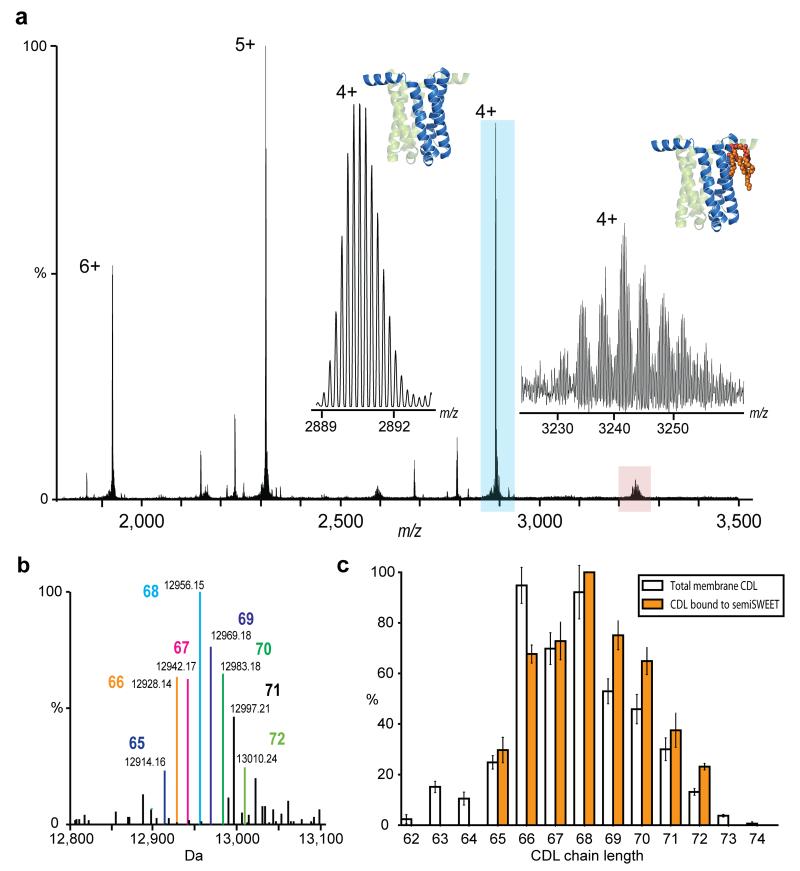
Examination of membrane protein semiSWEET in direct contact with bound lipids. (a) Isotopically resolved mass spectrum of semiSWEET released from DDM micelle obtained at an increased transient time of 256 ms. Monoisotopic mass measurement of the major protein peak (blue) gives an error of 0.9 ppm once N-terminal methionine cleavage had been taken into account. Inset shows a zoom of the apo protein peak (blue) and of the satellite peaks (red) revealing a distribution of homologous series of lipids. (b) Identification of directly bound lipids. Deconvoluted spectrum of semiSWEET with bound CDL adducts showing monoisotopic mass and enabling identification of lipid class as cardiolipin. Acyl chain length assignment is indicated. (c) Comparison of CDL bound to semiSWEET (orange bars) with the distribution of cardiolipin in the total membrane (clear bars). A clear shift of the distribution to longer acyl chain lengths for the lipids bound directly to semiSWEET can be discerned.
References
Methods Only References
-
- Hsu F-F, et al. Structural characterization of cardiolipin by tandem quadrupole and multiple-stage quadrupole ion-trap MS with electrospray ionization. Journal of the American Society for MS. 2005;16:491–504. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
