

TRPV1 function is modulated by Cdk5-mediated phosphorylation: insights into the molecular mechanism of nociception
- PMID: 26902776
- PMCID: PMC4763283
- DOI: 10.1038/srep22007
TRPV1 function is modulated by Cdk5-mediated phosphorylation: insights into the molecular mechanism of nociception
Abstract
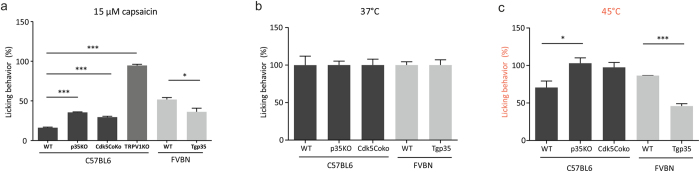
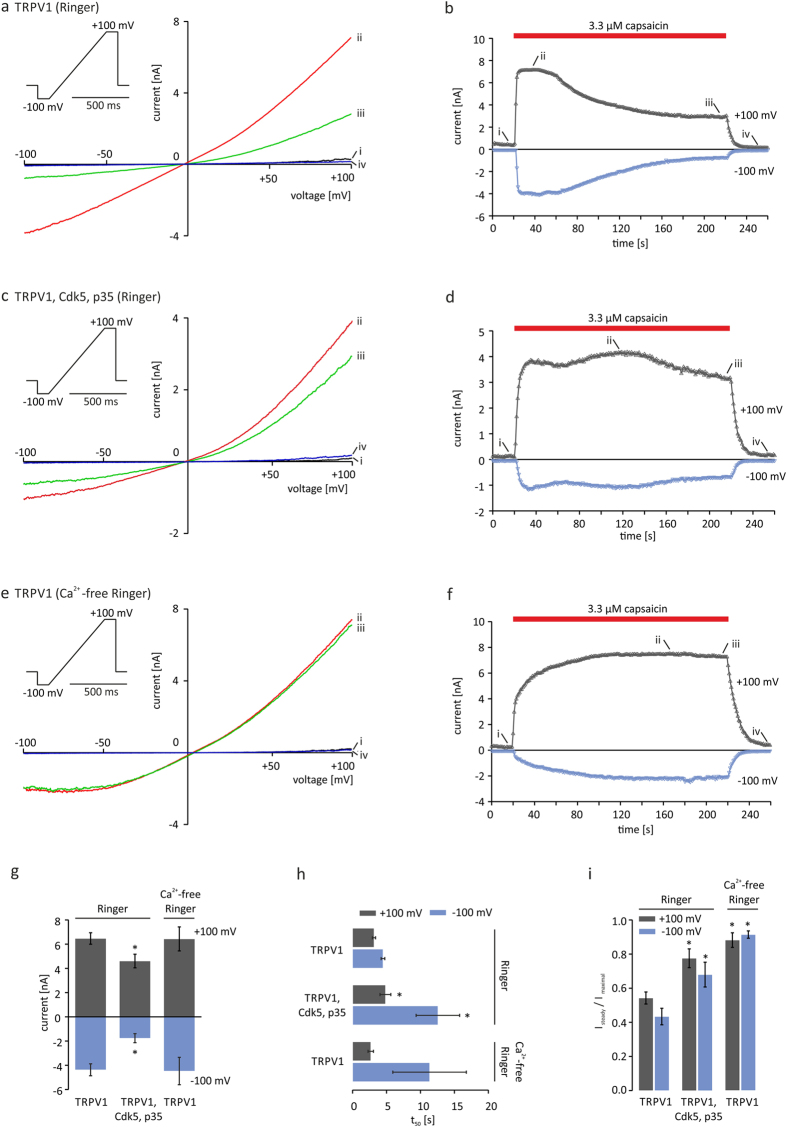
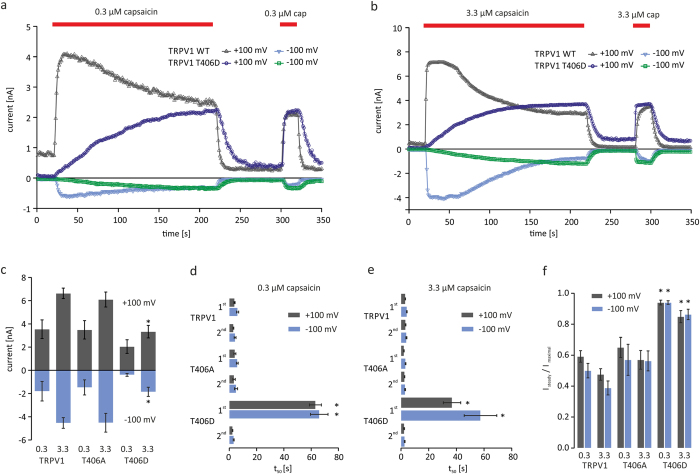
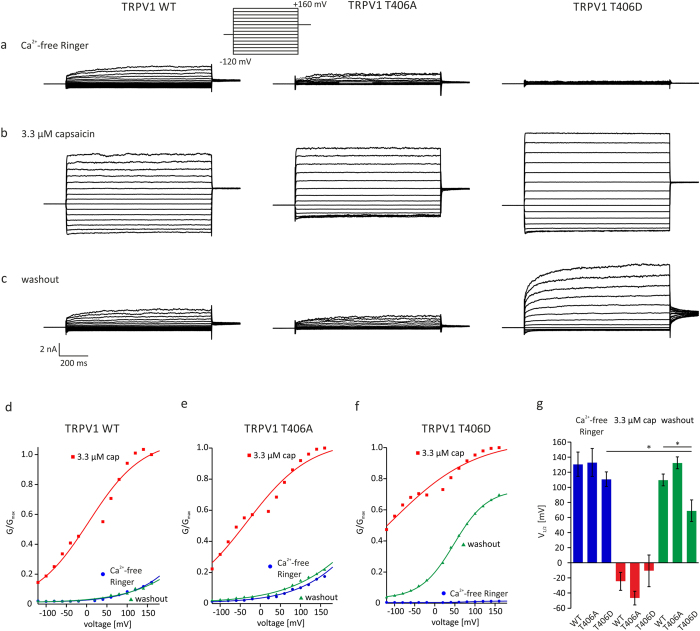
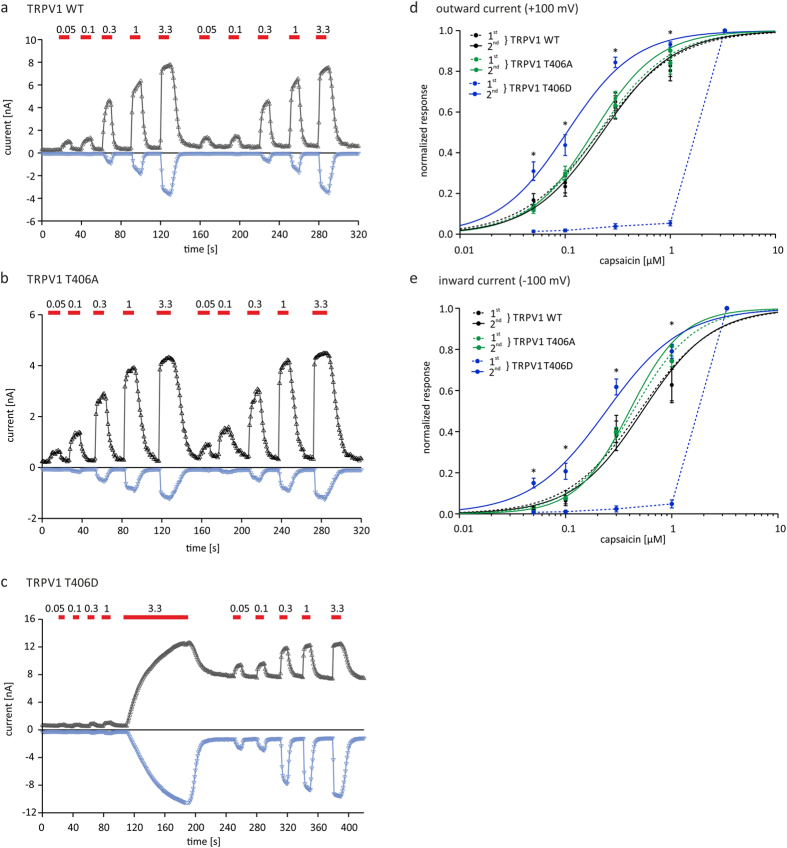
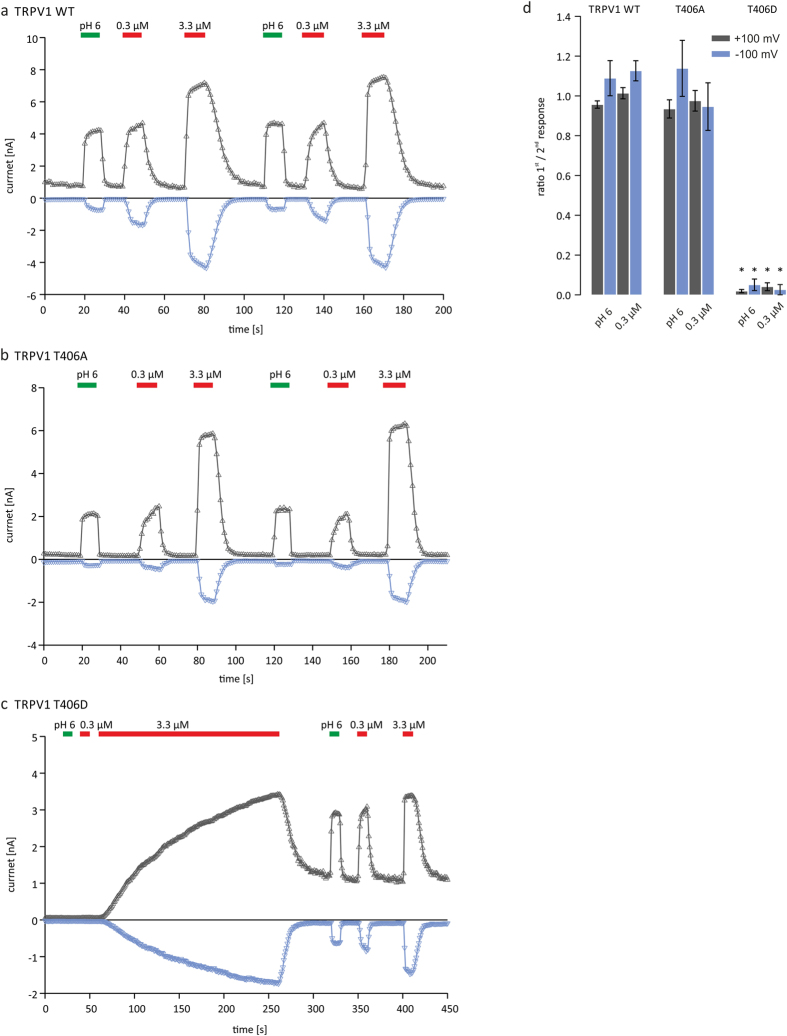
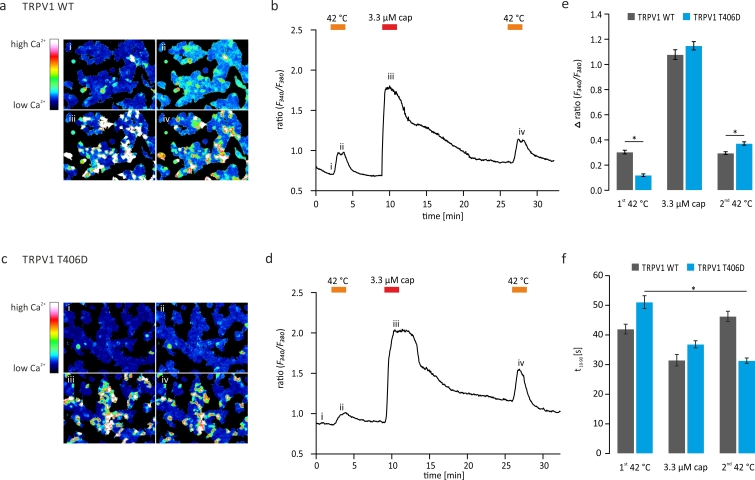
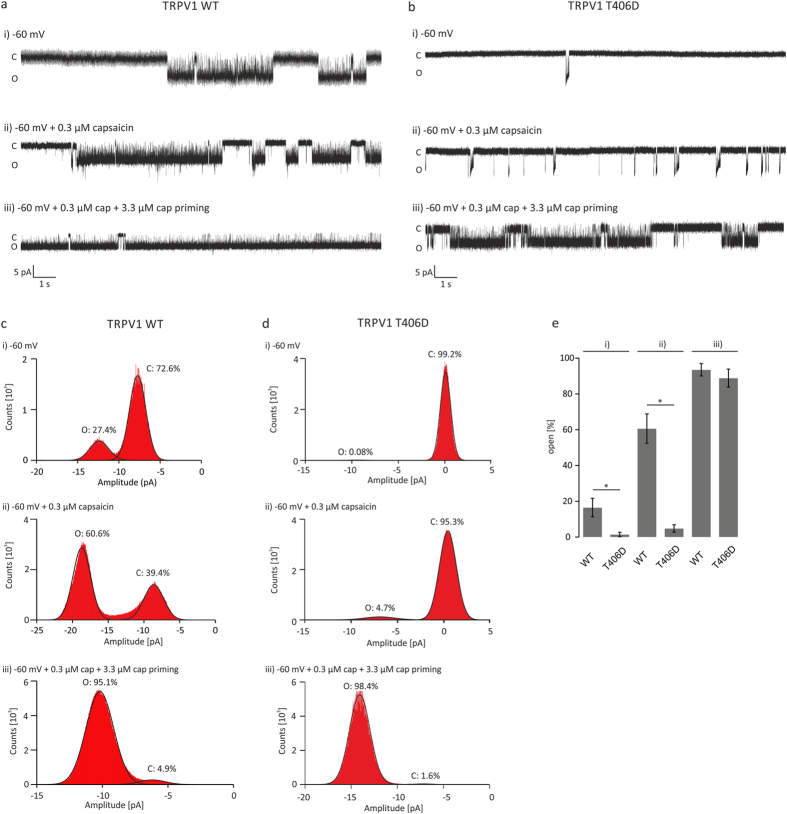
TRPV1 is a polymodally activated cation channel acting as key receptor in nociceptive neurons. Its function is strongly affected by kinase-mediated phosphorylation leading to hyperalgesia and allodynia. We present behavioral and molecular data indicating that TRPV1 is strongly modulated by Cdk5-mediated phosphorylation at position threonine-407(mouse)/T406(rat). Increasing or decreasing Cdk5 activity in genetically engineered mice has severe consequences on TRPV1-mediated pain perception leading to altered capsaicin consumption and sensitivity to heat. To understand the molecular and structural/functional consequences of TRPV1 phosphorylation, we generated various rTRPV1T406 receptor variants to mimic phosphorylated or dephosphorylated receptor protein. We performed detailed functional characterization by means of electrophysiological whole-cell and single-channel recordings as well as Ca(2+)-imaging and challenged recombinant rTRPV1 receptors with capsaicin, low pH, or heat. We found that position T406 is critical for the function of TRPV1 by modulating ligand-sensitivity, activation, and desensitization kinetics as well as voltage-dependence. Based on high resolution structures of TRPV1, we discuss T406 being involved in the molecular transition pathway, its phosphorylation leading to a conformational change and influencing the gating of the receptor. Cdk5-mediated phosphorylation of T406 can be regarded as an important molecular switch modulating TRPV1-related behavior and pain sensitivity.
Figures
References
-
- Szallasi A. & Blumberg P. M. Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol. Rev. 51, 159–212 (1999). - PubMed
-
- Caterina M. J. et al. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389, 816–824 (1997). - PubMed
-
- Zygmunt P. M. et al. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 400, 452–457 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
