

Vector-free transmission and persistence of Japanese encephalitis virus in pigs
- PMID: 26902924
- PMCID: PMC4766424
- DOI: 10.1038/ncomms10832
Vector-free transmission and persistence of Japanese encephalitis virus in pigs
Abstract
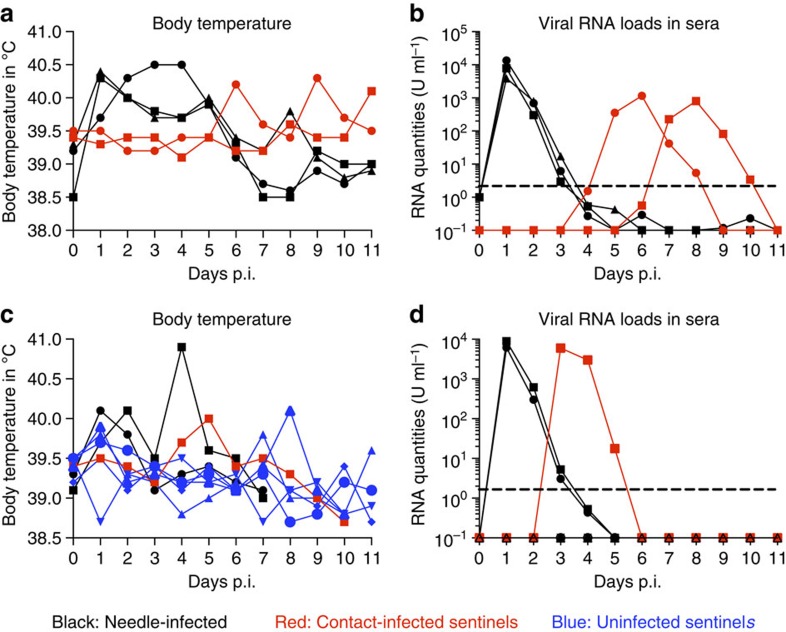
Japanese encephalitis virus (JEV), a main cause of severe viral encephalitis in humans, has a complex ecology, composed of a cycle involving primarily waterbirds and mosquitoes, as well as a cycle involving pigs as amplifying hosts. To date, JEV transmission has been exclusively described as being mosquito-mediated. Here we demonstrate that JEV can be transmitted between pigs in the absence of arthropod vectors. Pigs shed virus in oronasal secretions and are highly susceptible to oronasal infection. Clinical symptoms, virus tropism and central nervous system histological lesions are similar in pigs infected through needle, contact or oronasal inoculation. In all cases, a particularly important site of replication are the tonsils, in which JEV is found to persist for at least 25 days despite the presence of high levels of neutralizing antibodies. Our findings could have a major impact on the ecology of JEV in temperate regions with short mosquito seasons.
Figures




References
-
- Impoinvil D. E., Baylis M. & Solomon T. Japanese encephalitis: on the One Health agenda. Curr. Top. Microbiol. Immunol. 365, 205–247 (2013). - PubMed
-
- van den Hurk A. F., Ritchie S. A. & Mackenzie J. S. Ecology and geographical expansion of Japanese encephalitis virus. Annu. Rev. Entomol. 54, 17–35 (2009). - PubMed
