

Retinoic acid-induced premature osteoblast-to-preosteocyte transitioning has multiple effects on calvarial development
- PMID: 26903503
- PMCID: PMC4852491
- DOI: 10.1242/dev.129189
Retinoic acid-induced premature osteoblast-to-preosteocyte transitioning has multiple effects on calvarial development
Abstract
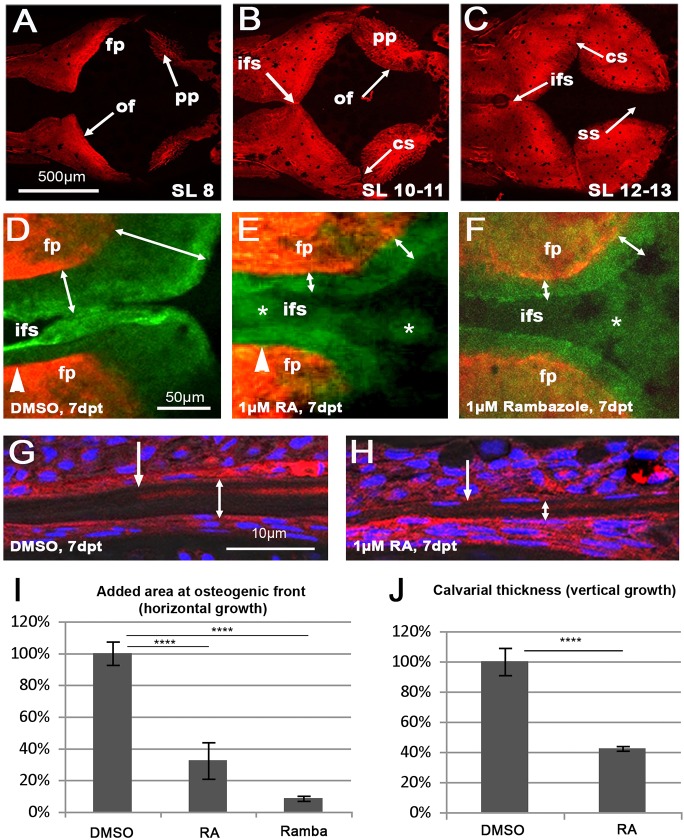
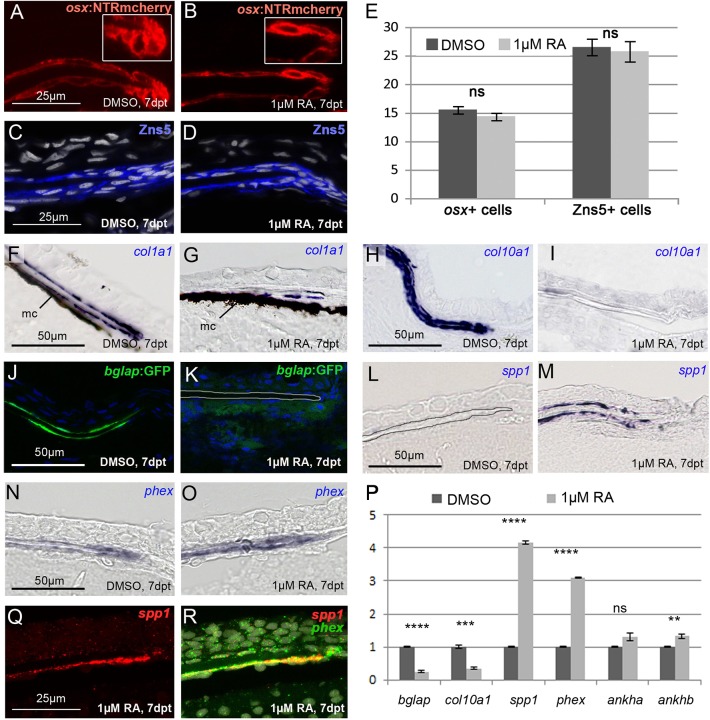
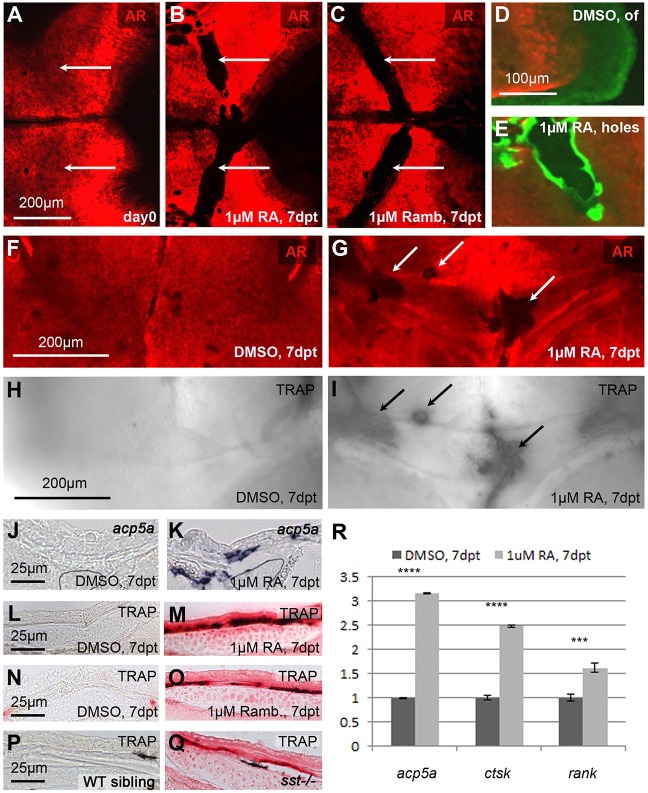
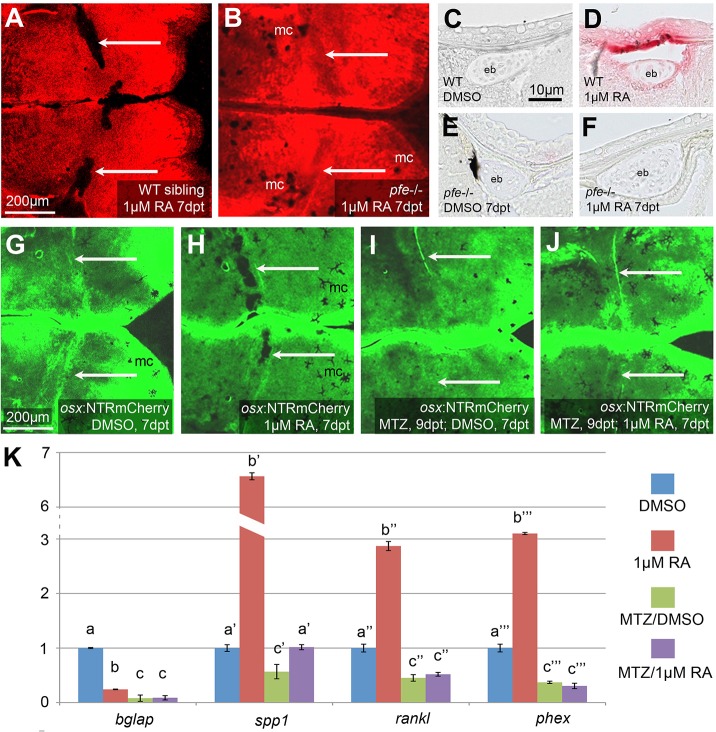
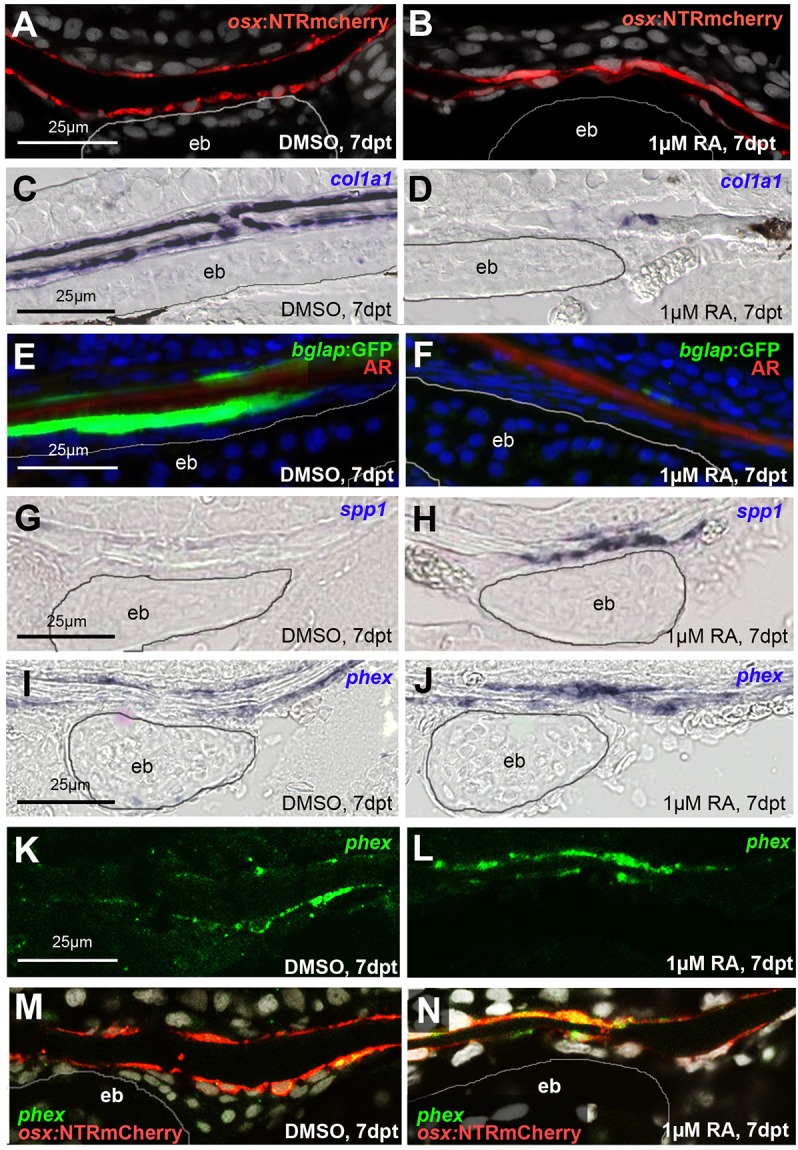
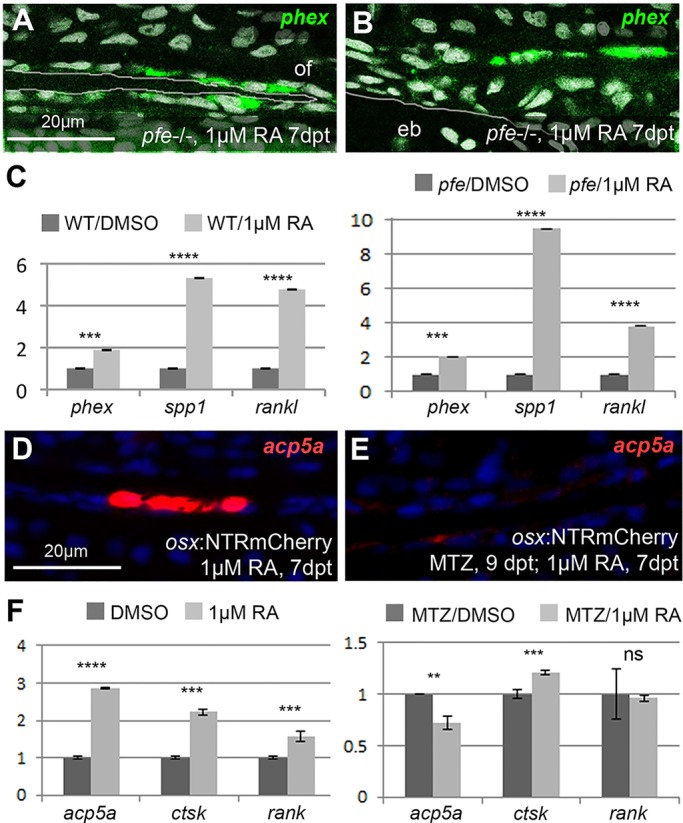
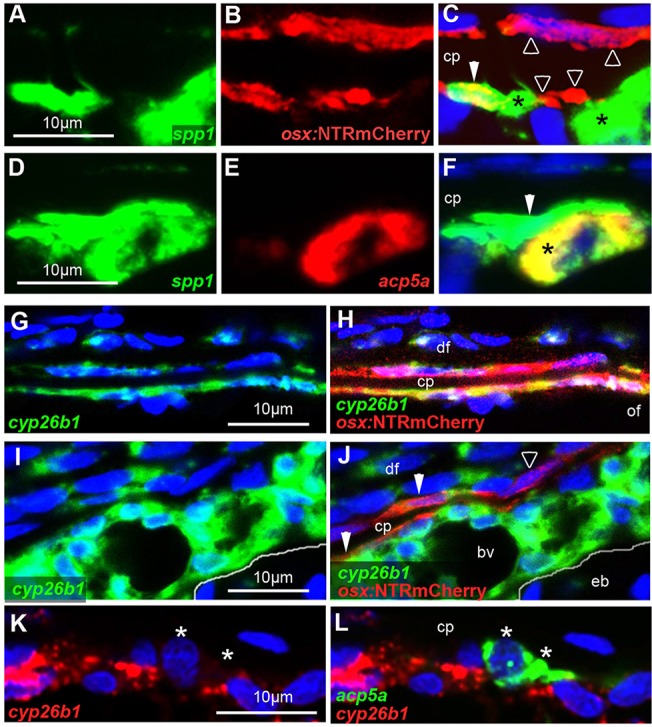
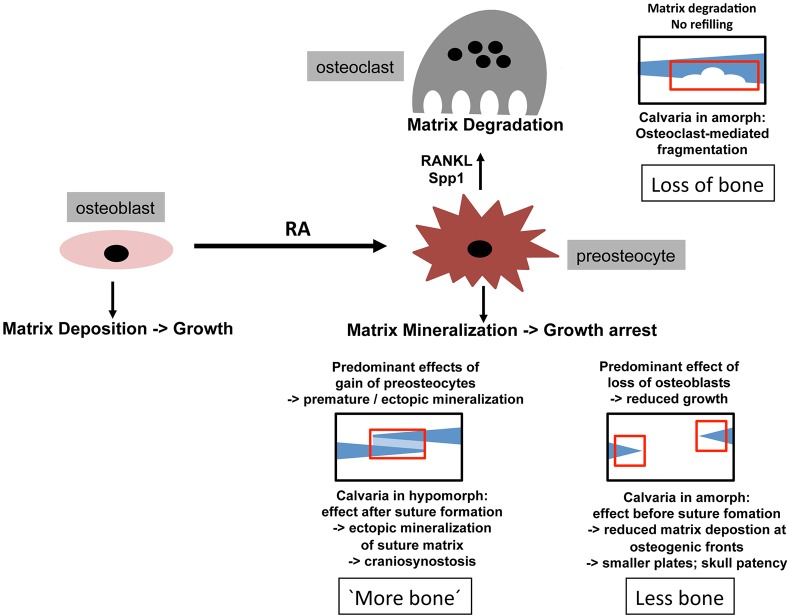
We have previously shown that, in human and zebrafish, hypomorphic mutations of the gene encoding the retinoic acid (RA)-metabolizing enzyme Cyp26b1 result in coronal craniosynostosis, caused by an RA-induced premature transitioning of suture osteoblasts to preosteocytes, inducing ectopic mineralization of the suture's osteoid matrix. In addition, we showed that human CYP26B1 null patients have more severe and seemingly opposite skull defects, characterized by smaller and fragmented calvaria, but the cellular basis of these defects remained largely unclear. Here, by treating juvenile zebrafish with exogenous RA or a chemical Cyp26 inhibitor in the presence or absence of osteogenic cells or bone-resorbing osteoclasts, we demonstrate that both reduced calvarial size and calvarial fragmentation are also caused by RA-induced premature osteoblast-to-preosteocyte transitioning. During calvarial growth, the resulting osteoblast deprival leads to decreased osteoid production and thereby smaller and thinner calvaria, whereas calvarial fragmentation is caused by increased osteoclast stimulation through the gained preosteocytes. Together, our data demonstrate that RA-induced osteoblast-to-preosteocyte transitioning has multiple effects on developing bone in Cyp26b1 mutants, ranging from gain to loss of bone, depending on the allelic strength, the developmental stage and the cellular context.
Keywords: Bone development; Diversity; Retinoic acid; Zebrafish.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
Similar articles
-
Craniosynostosis and multiple skeletal anomalies in humans and zebrafish result from a defect in the localized degradation of retinoic acid.Am J Hum Genet. 2011 Nov 11;89(5):595-606. doi: 10.1016/j.ajhg.2011.09.015. Epub 2011 Oct 20. Am J Hum Genet. 2011. PMID: 22019272 Free PMC article.
-
Restriction of retinoic acid activity by Cyp26b1 is required for proper timing and patterning of osteogenesis during zebrafish development.Development. 2008 Nov;135(22):3775-87. doi: 10.1242/dev.021238. Epub 2008 Oct 16. Development. 2008. PMID: 18927157 Free PMC article.
-
Retinoic acid and Cyp26b1 are critical regulators of osteogenesis in the axial skeleton.Development. 2008 Nov;135(22):3765-74. doi: 10.1242/dev.024034. Epub 2008 Oct 16. Development. 2008. PMID: 18927155
-
Ets transcription factors and targets in osteogenesis.Oncogene. 2000 Dec 18;19(55):6455-63. doi: 10.1038/sj.onc.1204037. Oncogene. 2000. PMID: 11175361 Review.
-
Buried alive: how osteoblasts become osteocytes.Dev Dyn. 2006 Jan;235(1):176-90. doi: 10.1002/dvdy.20603. Dev Dyn. 2006. PMID: 16258960 Review.
Cited by
-
Role of vitamins beyond vitamin D3 in bone health and osteoporosis (Review).Int J Mol Med. 2024 Jan;53(1):9. doi: 10.3892/ijmm.2023.5333. Epub 2023 Dec 8. Int J Mol Med. 2024. PMID: 38063255 Free PMC article. Review.
-
Osteocytogenesis: Roles of Physicochemical Factors, Collagen Cleavage, and Exogenous Molecules.Tissue Eng Part B Rev. 2018 Jun;24(3):215-225. doi: 10.1089/ten.teb.2017.0378. Epub 2018 Jan 5. Tissue Eng Part B Rev. 2018. PMID: 29304315 Free PMC article. Review.
-
A role for G protein-coupled receptor 137b in bone remodeling in mouse and zebrafish.Bone. 2019 Oct;127:104-113. doi: 10.1016/j.bone.2019.06.002. Epub 2019 Jun 5. Bone. 2019. PMID: 31173907 Free PMC article.
-
Excessive dietary intake of vitamin A reduces skull bone thickness in mice.PLoS One. 2017 Apr 20;12(4):e0176217. doi: 10.1371/journal.pone.0176217. eCollection 2017. PLoS One. 2017. PMID: 28426756 Free PMC article.
-
Direct BMP signaling to chordoblasts is required for the initiation of segmented notochord sheath mineralization in zebrafish vertebral column development.Front Endocrinol (Lausanne). 2023 May 8;14:1107339. doi: 10.3389/fendo.2023.1107339. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37223044 Free PMC article.
References
-
- Apschner A., Huitema L. F. A., Ponsioen B., Peterson-Maduro J. and Schulte-Merker S. (2014). Zebrafish enpp1 mutants exhibit pathological mineralization, mimicking features of generalized arterial calcification of infancy (GACI) and pseudoxanthoma elasticum (PXE). Dis. Model. Mech. 7, 811-822. 10.1242/dmm.015693 - DOI - PMC - PubMed
-
- Barros N. M. T., Hoac B., Neves R. L., Addison W. N., Assis D. M., Murshed M., Carmona A. K. and McKee M. D. (2013). Proteolytic processing of osteopontin by PHEX and accumulation of osteopontin fragments in Hyp mouse bone, the murine model of X-linked hypophosphatemia. J. Bone Miner. Res. 28, 688-699. 10.1002/jbmr.1766 - DOI - PubMed
-
- Begemann G., Schilling T. F., Rauch G. J., Geisler R. and Ingham P. W. (2001). The zebrafish neckless mutation reveals a requirement for raldh2 in mesodermal signals that pattern the hindbrain. Development 128, 3081-3094. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
