

Major Histocompatibility Complex (MHC) Class I Processing of the NY-ESO-1 Antigen Is Regulated by Rpn10 and Rpn13 Proteins and Immunoproteasomes following Non-lysine Ubiquitination
- PMID: 26903513
- PMCID: PMC4861448
- DOI: 10.1074/jbc.M115.705178
Major Histocompatibility Complex (MHC) Class I Processing of the NY-ESO-1 Antigen Is Regulated by Rpn10 and Rpn13 Proteins and Immunoproteasomes following Non-lysine Ubiquitination
Abstract
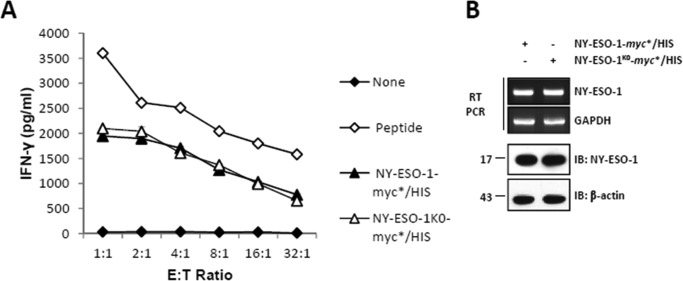
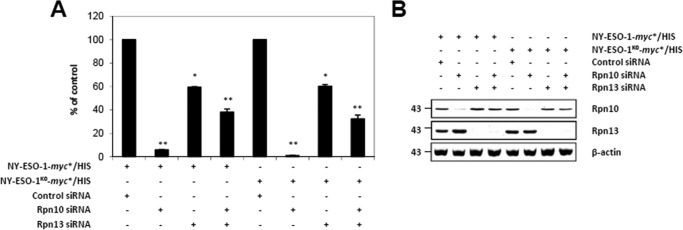
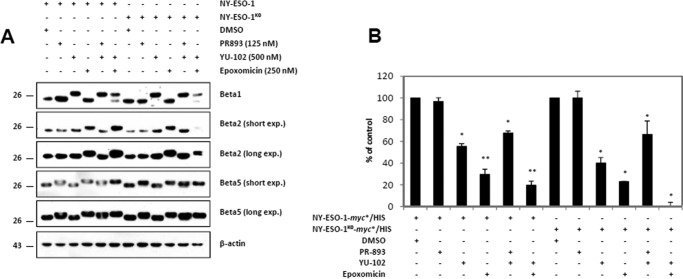
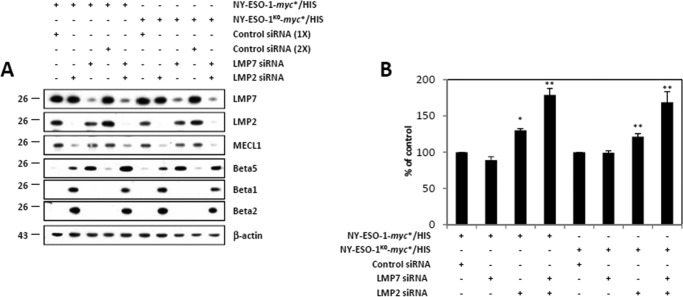
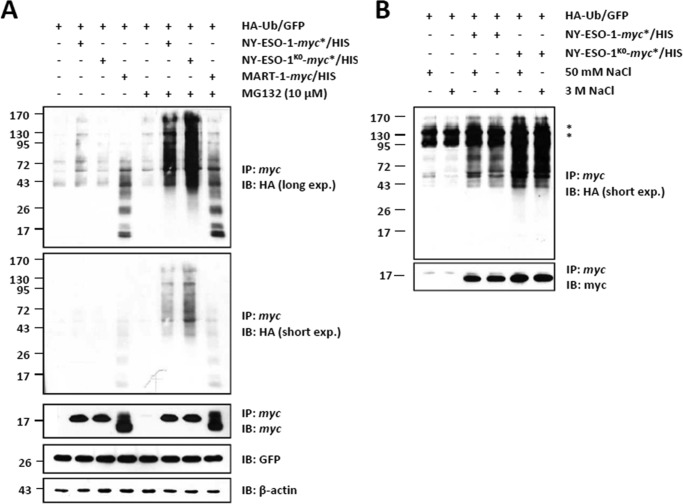
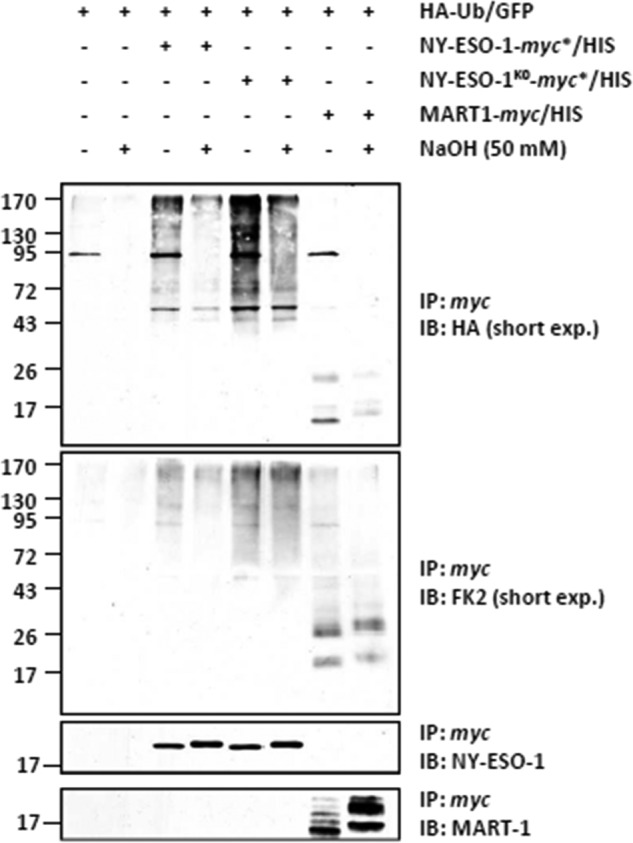
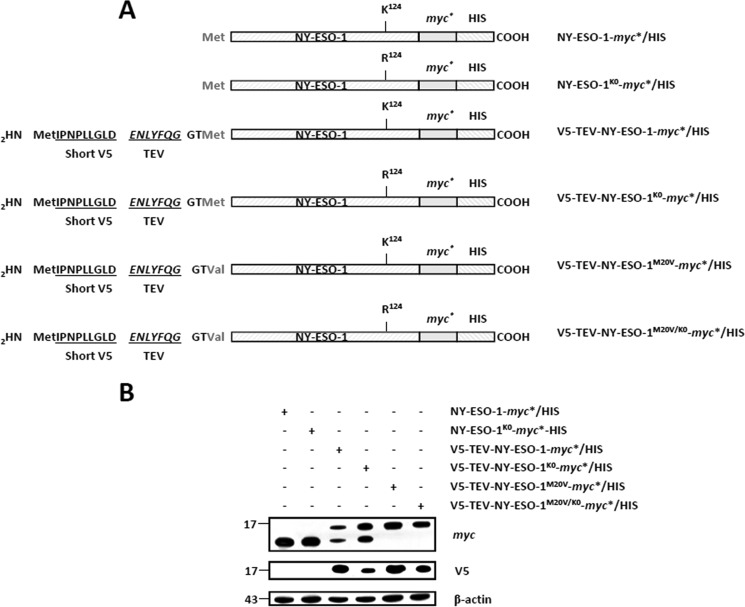
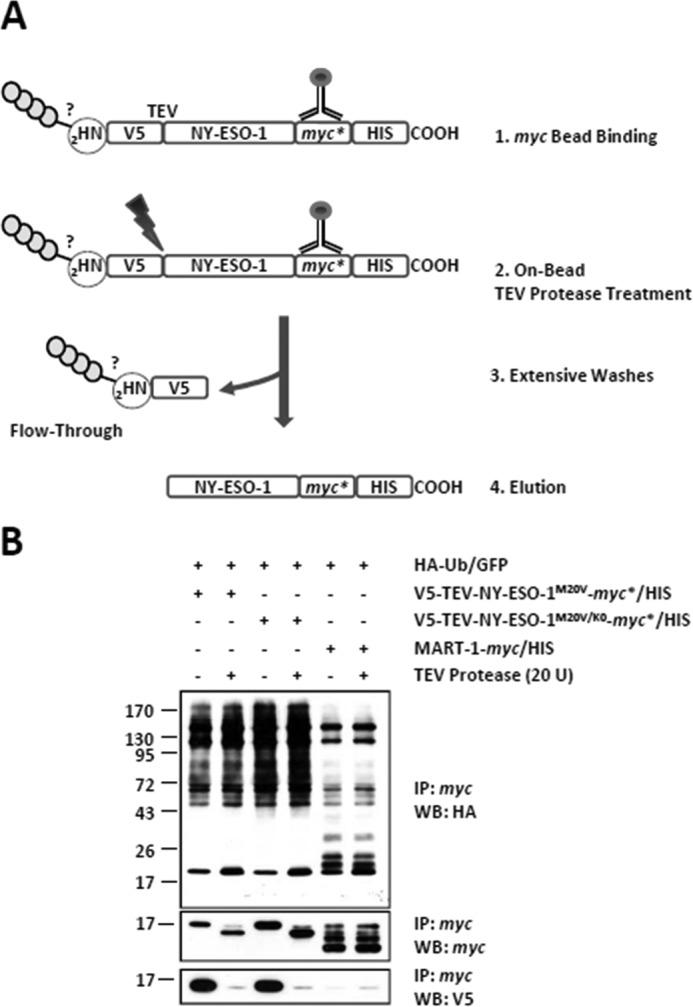
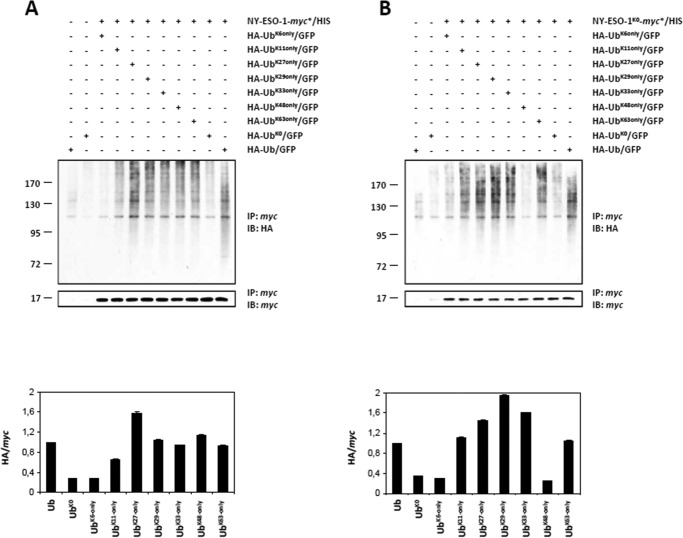
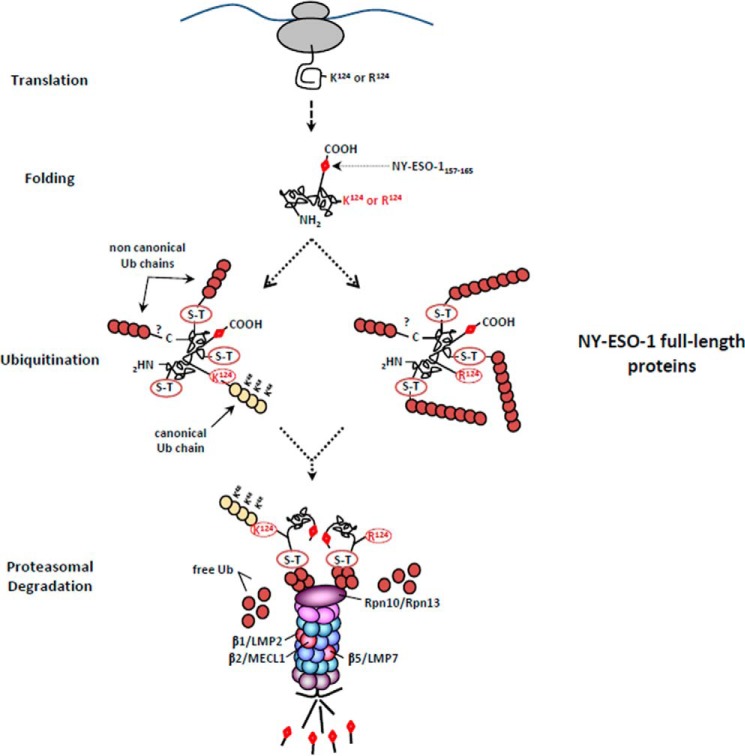
The supply of MHC class I-restricted peptides is primarily ensured by the degradation of intracellular proteins via the ubiquitin-proteasome system. Depending on the target and the enzymes involved, ubiquitination is a process that may dramatically vary in terms of linkages, length, and attachment sites. Here we identified the unique lysine residue at position 124 of the NY-ESO-1 cancer/testis antigen as the acceptor site for the formation of canonical Lys-48-linkages. Interestingly, a lysine-less form of NY-ESO-1 was as efficient as its wild-type counterpart in supplying the HLA-A*0201-restricted NY-ESO-1157-165 antigenic peptide. In fact, we show that the regulation of NY-ESO-1 processing by the ubiquitin receptors Rpn10 and Rpn13 as a well as by the standard and immunoproteasome is governed by non-canonical ubiquitination on non-lysine sites. In summary, our data underscore the significance of atypical ubiquitination in the modulation of MHC class I antigen processing.
Keywords: antigen processing; proteasome; protein degradation; tumor immunology; ubiquitin.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Rock K. L., York I. A., Saric T., and Goldberg A. L. (2002) Protein degradation and the generation of MHC class I-presented peptides. Adv. Immunol. 80, 1–70 - PubMed
-
- Murata S., Yashiroda H., and Tanaka K. (2009) Molecular mechanisms of proteasome assembly. Nat. Rev. Mol. Cell Biol. 10, 104–115 - PubMed
-
- Ciechanover A., and Stanhill A. (2014) The complexity of recognition of ubiquitinated substrates by the 26S proteasome. Biochim. Biophys. Acta 1843, 86–96 - PubMed
-
- DeMartino G. N., and Slaughter C. A. (1999) The proteasome, a novel protease regulated by multiple mechanisms. J. Biol. Chem. 274, 22123–22126 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
