

Tetratricopeptide repeat protein protects photosystem I from oxidative disruption during assembly
- PMID: 26903622
- PMCID: PMC4791029
- DOI: 10.1073/pnas.1524040113
Tetratricopeptide repeat protein protects photosystem I from oxidative disruption during assembly
Abstract
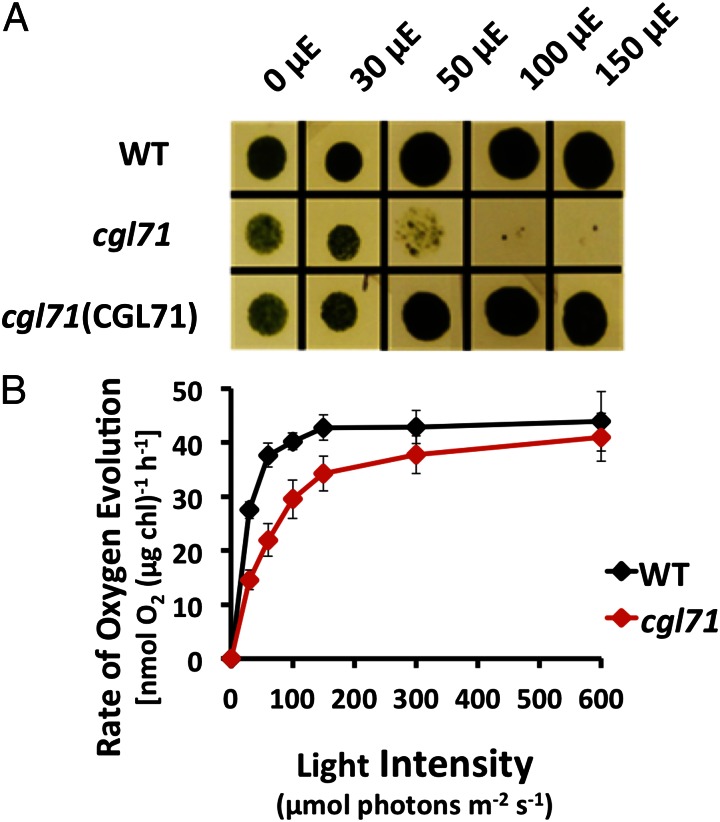
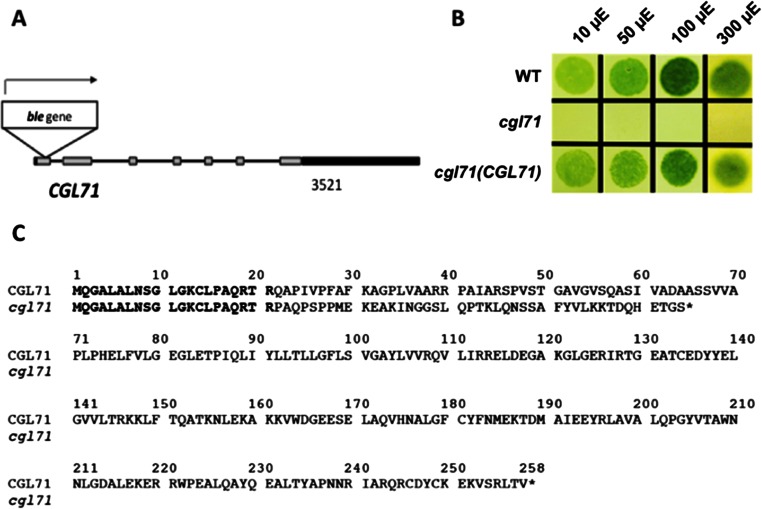
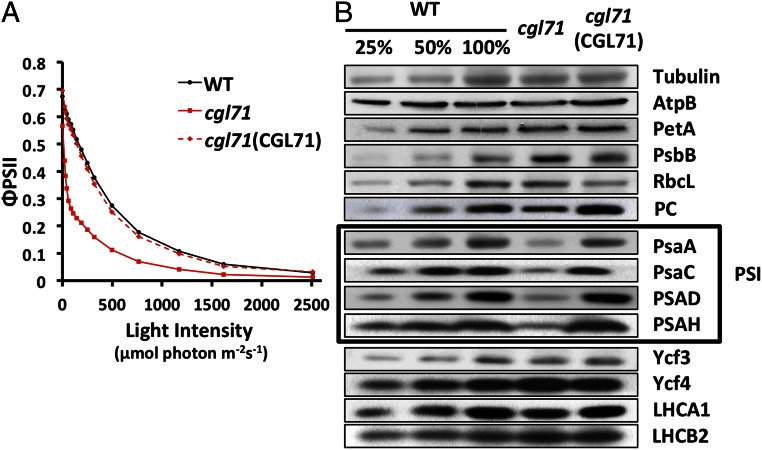
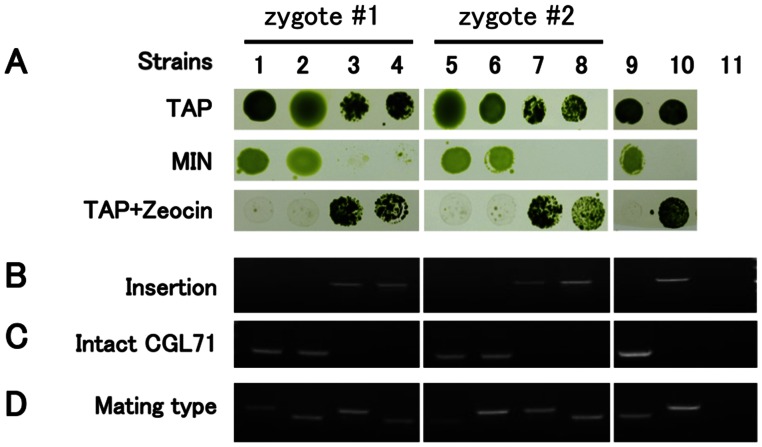
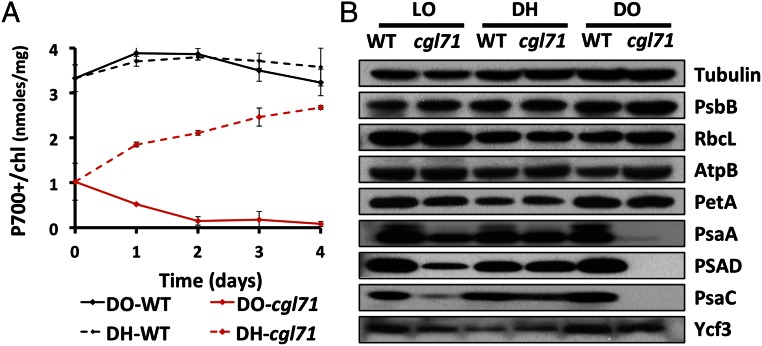
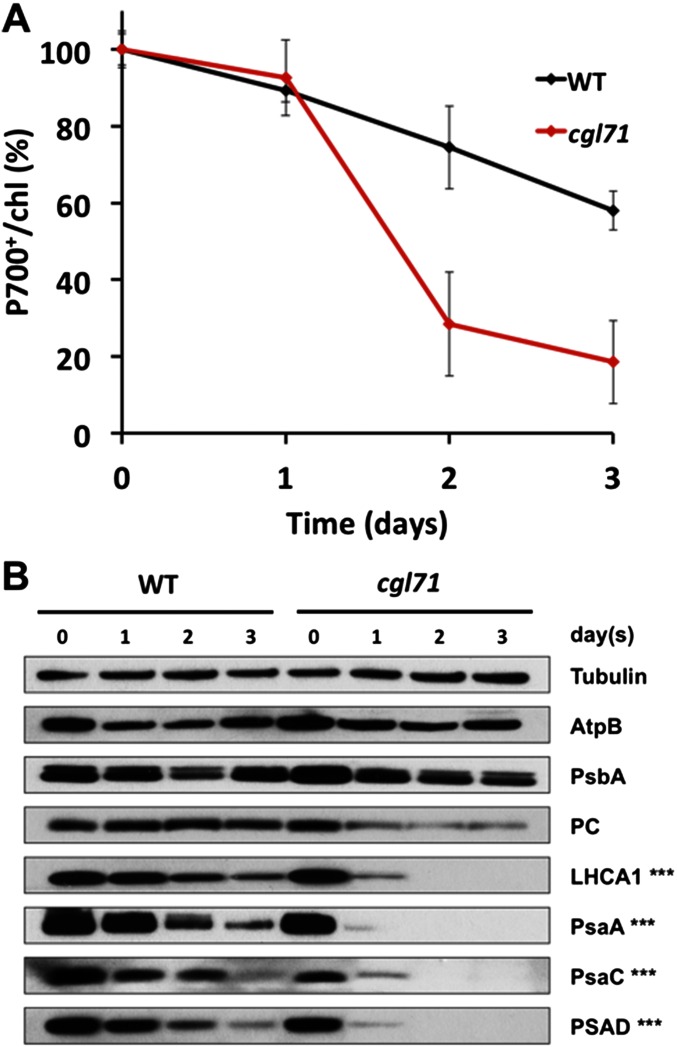
A Chlamydomonas reinhardtii mutant lacking CGL71, a thylakoid membrane protein previously shown to be involved in photosystem I (PSI) accumulation, exhibited photosensitivity and highly reduced abundance of PSI under photoheterotrophic conditions. Remarkably, the PSI content of this mutant declined to nearly undetectable levels under dark, oxic conditions, demonstrating that reduced PSI accumulation in the mutant is not strictly the result of photodamage. Furthermore, PSI returns to nearly wild-type levels when the O2 concentration in the medium is lowered. Overall, our results suggest that the accumulation of PSI in the mutant correlates with the redox state of the stroma rather than photodamage and that CGL71 functions under atmospheric O2 conditions to allow stable assembly of PSI. These findings may reflect the history of the Earth's atmosphere as it transitioned from anoxic to highly oxic (1-2 billion years ago), a change that required organisms to evolve mechanisms to assist in the assembly and stability of proteins or complexes with O2-sensitive cofactors.
Keywords: GreenCut; oxidative disruption; photosynthesis; photosystem I biogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
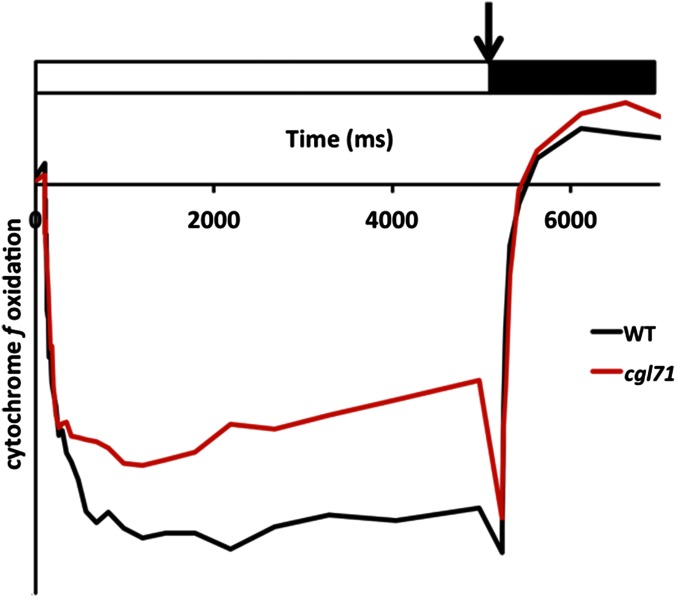
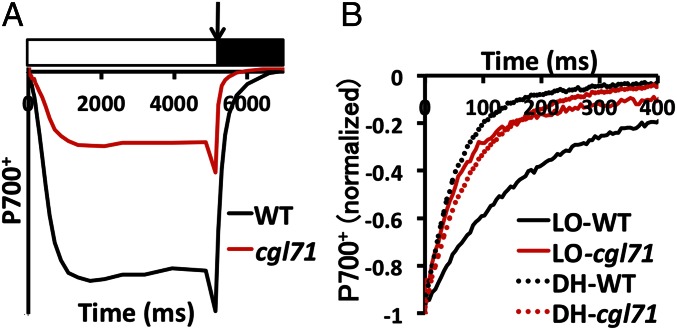
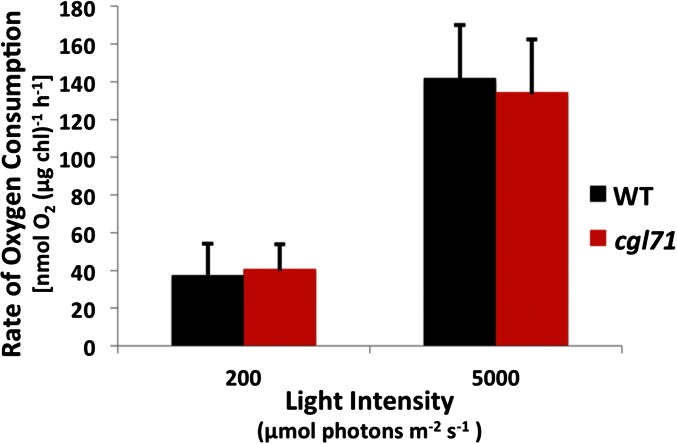
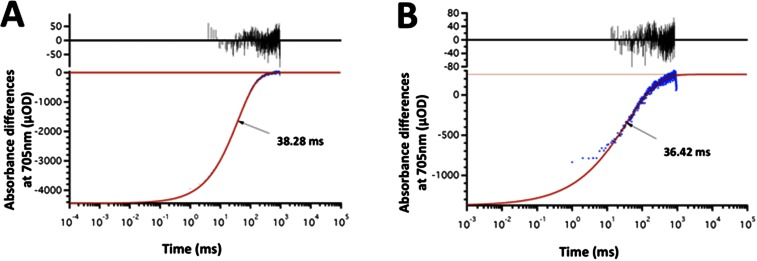

References
-
- Golbeck JH. Structure and function of photosystem I. Annu Rev Plant Physiol Plant Mol Biol. 1992;43:293–324.
-
- Chitnis PR. PHOTOSYSTEM I: Function and physiology. Annu Rev Plant Physiol Plant Mol Biol. 2001;52:593–626. - PubMed
-
- Shikanai T. Cyclic electron transport around photosystem I: Genetic approaches. Annu Rev Plant Biol. 2007;58:199–217. - PubMed
-
- Busch A, Hippler M. The structure and function of eukaryotic photosystem I. Biochim Biophys Acta. 2011;1807(8):864–877. - PubMed
-
- Kargul J, Janna Olmos JD, Krupnik T. Structure and function of photosystem I and its application in biomimetic solar-to-fuel systems. J Plant Physiol. 2012;169(16):1639–1653. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
