

c21orf59/kurly Controls Both Cilia Motility and Polarization
- PMID: 26904945
- PMCID: PMC4775428
- DOI: 10.1016/j.celrep.2016.01.069
c21orf59/kurly Controls Both Cilia Motility and Polarization
Abstract
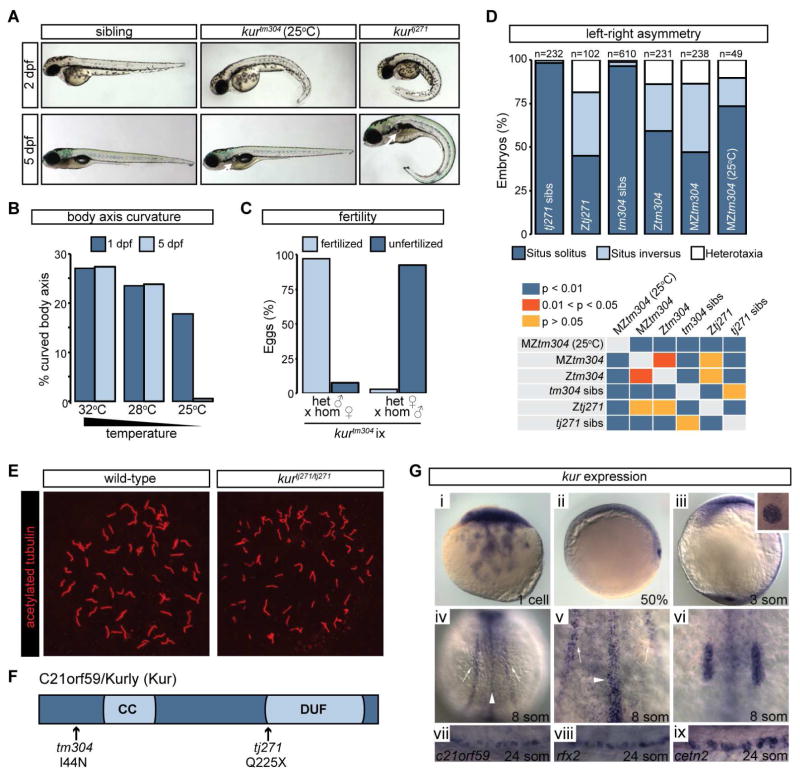
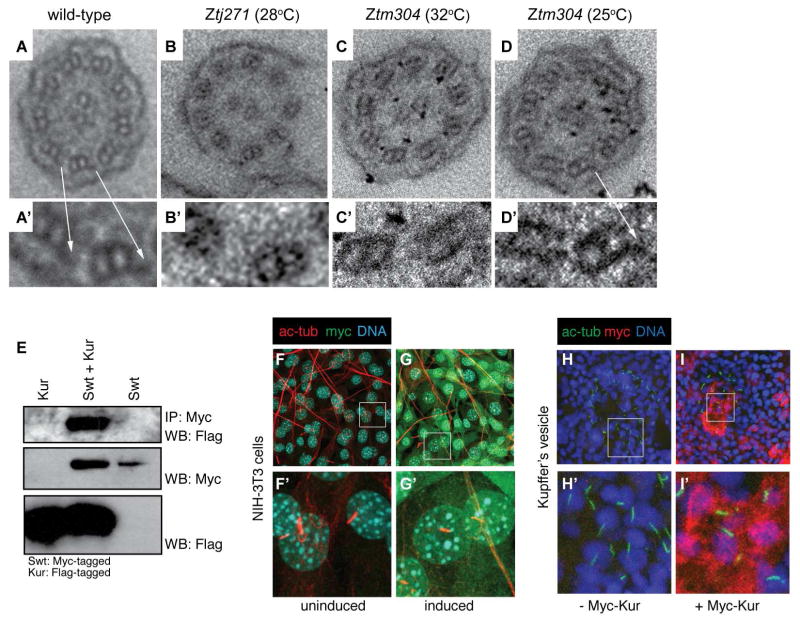
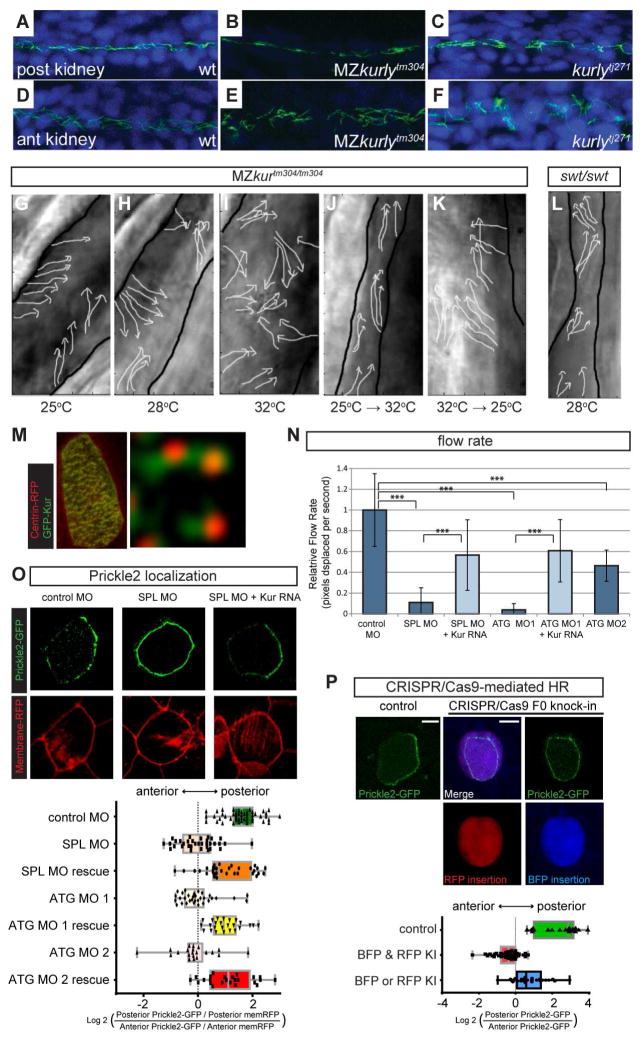
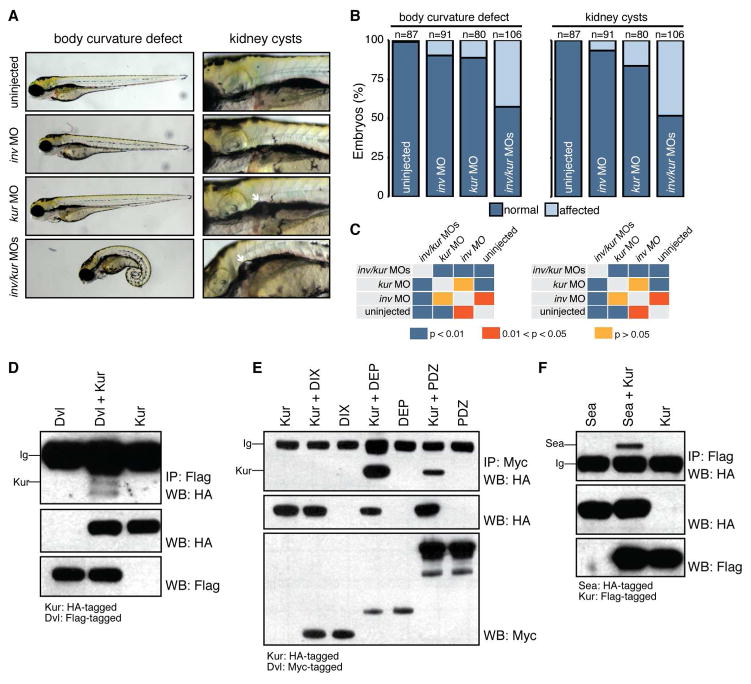
Cilia are microtubule-based projections that function in the movement of extracellular fluid. This requires cilia to be: (1) motile and driven by dynein complexes and (2) correctly polarized on the surface of cells, which requires planar cell polarity (PCP). Few factors that regulate both processes have been discovered. We reveal that C21orf59/Kurly (Kur), a cytoplasmic protein with some enrichment at the base of cilia, is needed for motility; zebrafish mutants exhibit characteristic developmental abnormalities and dynein arm defects. kur was also required for proper cilia polarization in the zebrafish kidney and the larval skin of Xenopus laevis. CRISPR/Cas9 coupled with homologous recombination to disrupt the endogenous kur locus in Xenopus resulted in the asymmetric localization of the PCP protein Prickle2 being lost in mutant multiciliated cells. Kur also makes interactions with other PCP components, including Disheveled. This supports a model wherein Kur plays a dual role in cilia motility and polarization.
Keywords: Kurly (Kur); c21orf59; cilia; ciliopathy; disheveled; multiciliated cell; planar cell polarity; primary ciliary dyskinesia.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Auer TO, Del Bene F. CRISPR/Cas9 and TALEN-mediated knock-in approaches in zebrafish. Methods. 2014;69:142–150. - PubMed
-
- Austin-Tse C, Halbritter J, Zariwala MA, Gilberti RM, Gee HY, Hellman N, Pathak N, Liu Y, Panizzi JR, Patel-King RS, et al. Zebrafish Ciliopathy Screen Plus Human Mutational Analysis Identifies C21orf59 and CCDC65 Defects as Causing Primary Ciliary Dyskinesia. American journal of human genetics. 2013;93:672–686. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
