

PP2A as a master regulator of the cell cycle
- PMID: 26906453
- PMCID: PMC4905575
- DOI: 10.3109/10409238.2016.1143913
PP2A as a master regulator of the cell cycle
Abstract
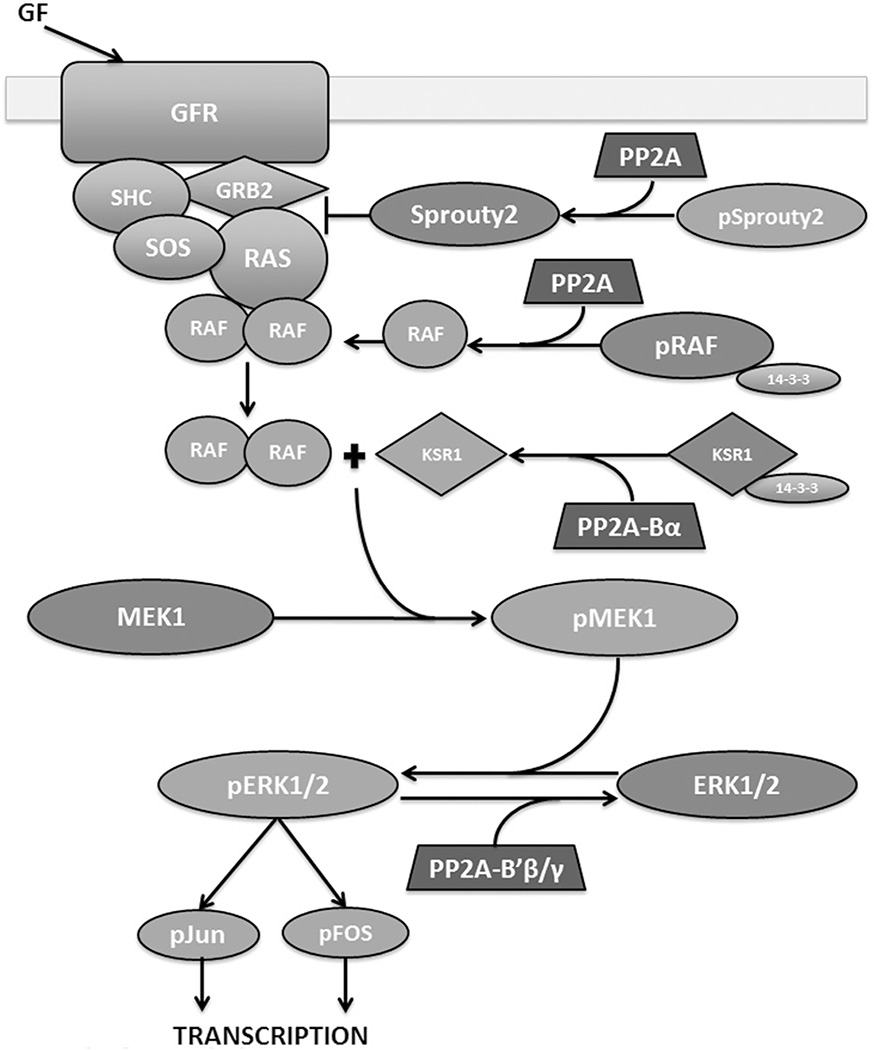
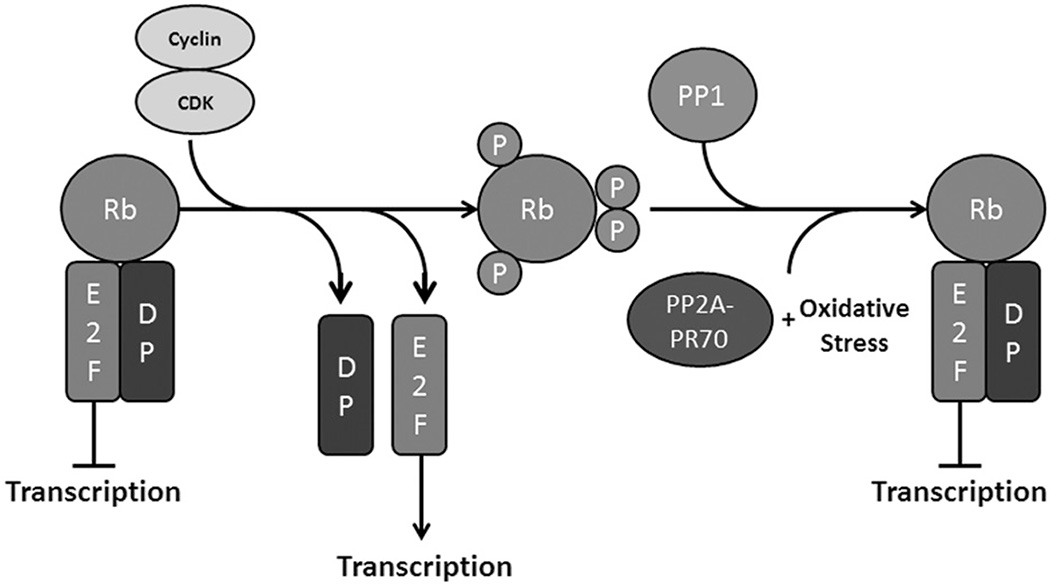
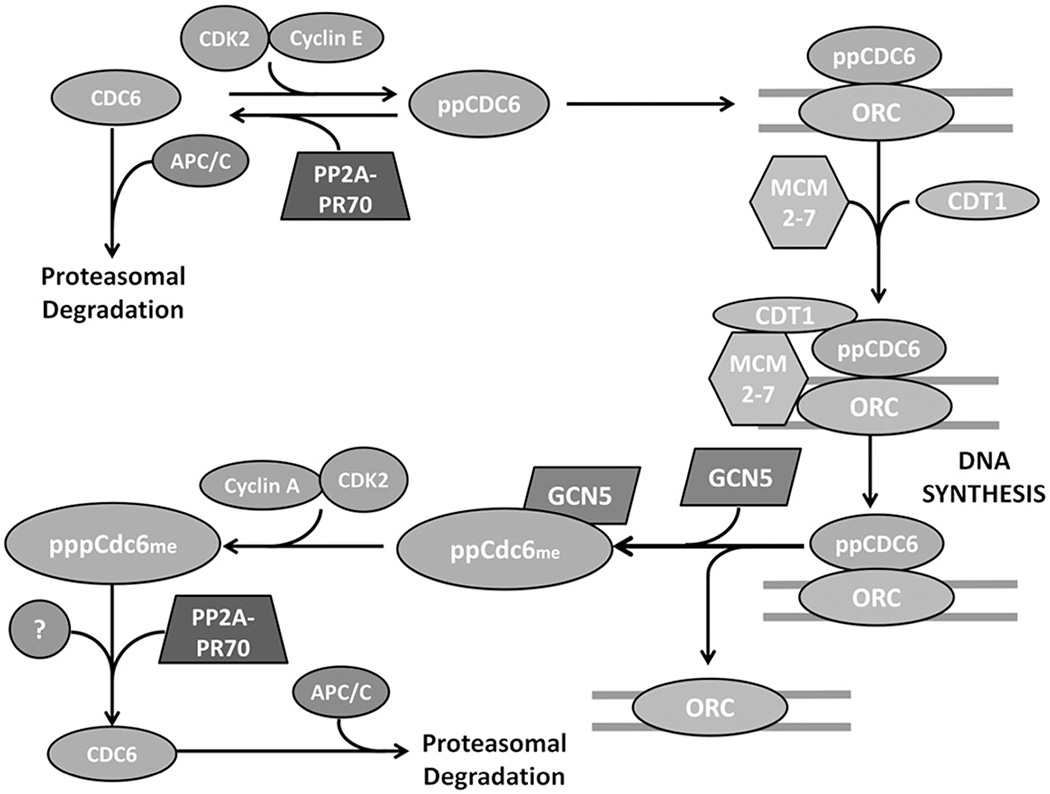
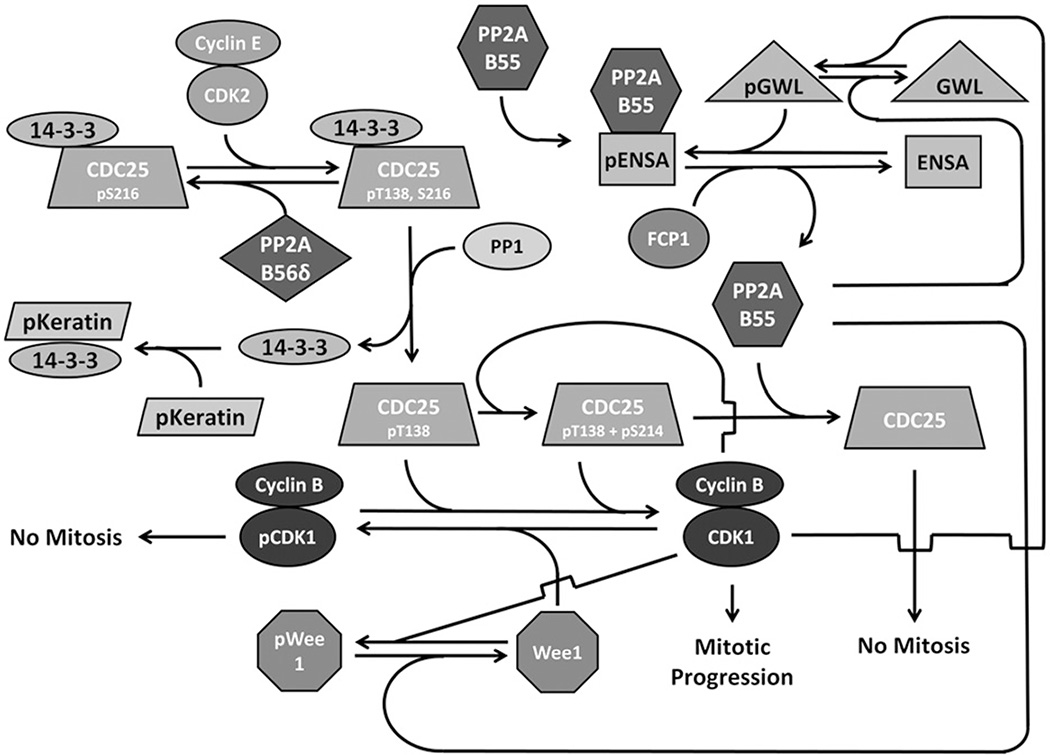
Protein phosphatase 2A (PP2A) plays a critical multi-faceted role in the regulation of the cell cycle. It is known to dephosphorylate over 300 substrates involved in the cell cycle, regulating almost all major pathways and cell cycle checkpoints. PP2A is involved in such diverse processes by the formation of structurally distinct families of holoenzymes, which are regulated spatially and temporally by specific regulators. Here, we review the involvement of PP2A in the regulation of three cell signaling pathways: wnt, mTOR and MAP kinase, as well as the G1→S transition, DNA synthesis and mitotic initiation. These processes are all crucial for proper cell survival and proliferation and are often deregulated in cancer and other diseases.
Keywords: Cancer; PP2A; cell cycle; cell division; mitosis; phosphatase.
Figures
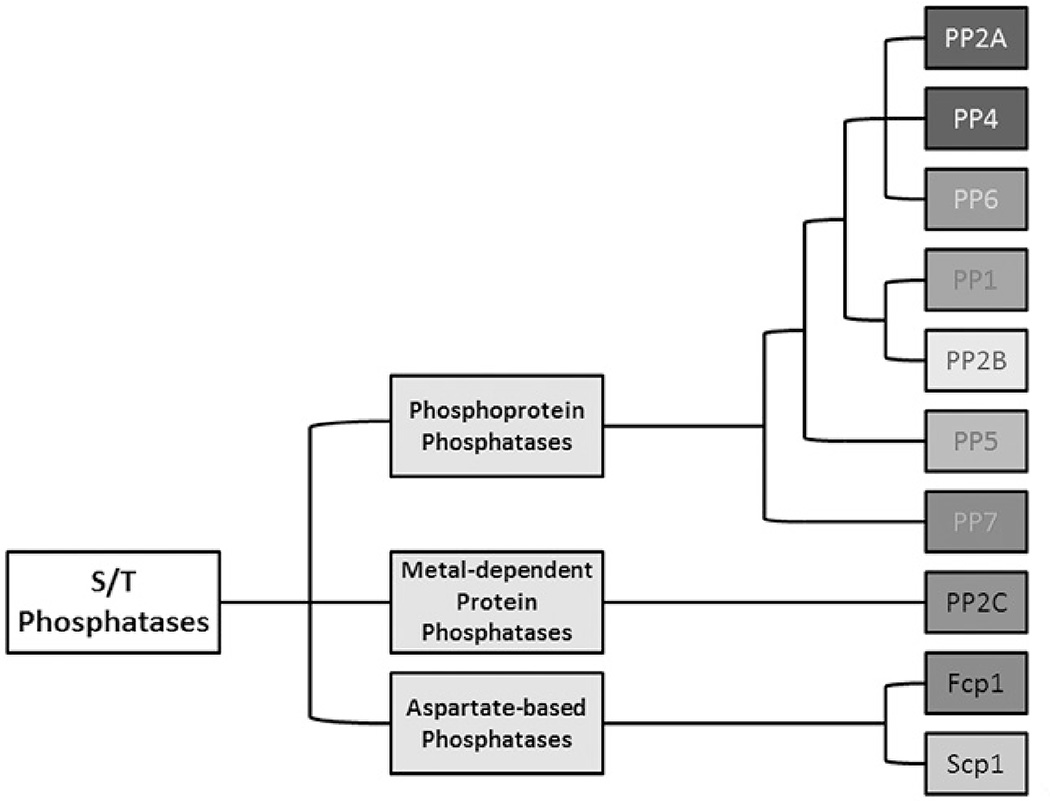
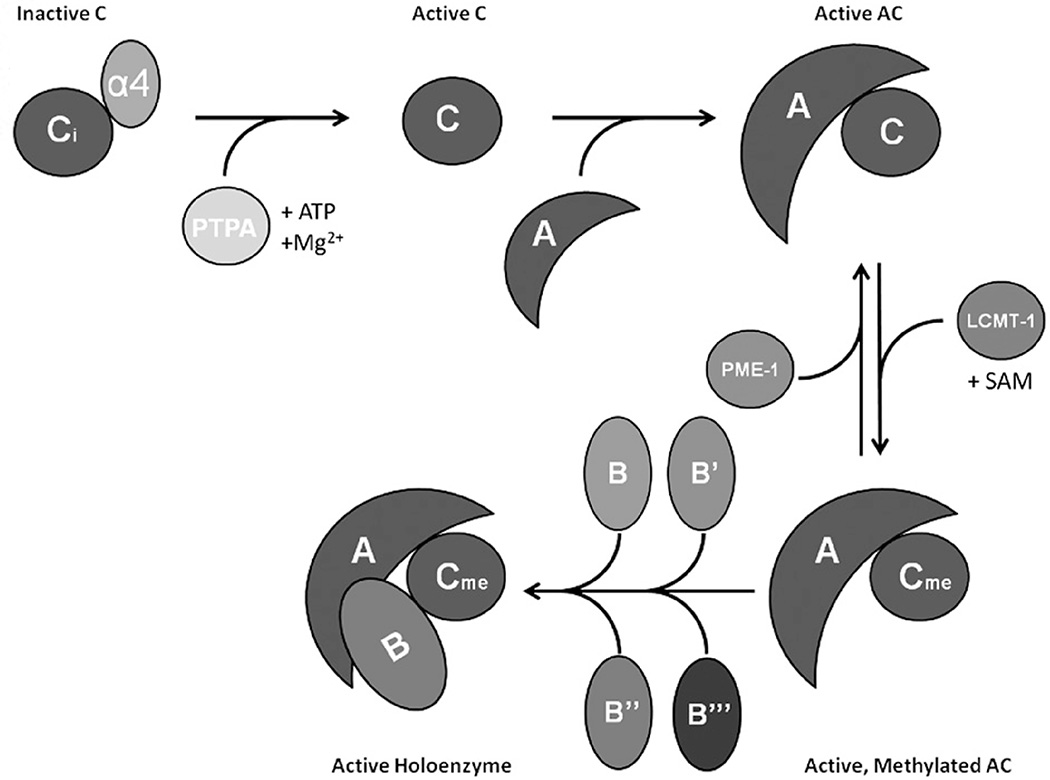
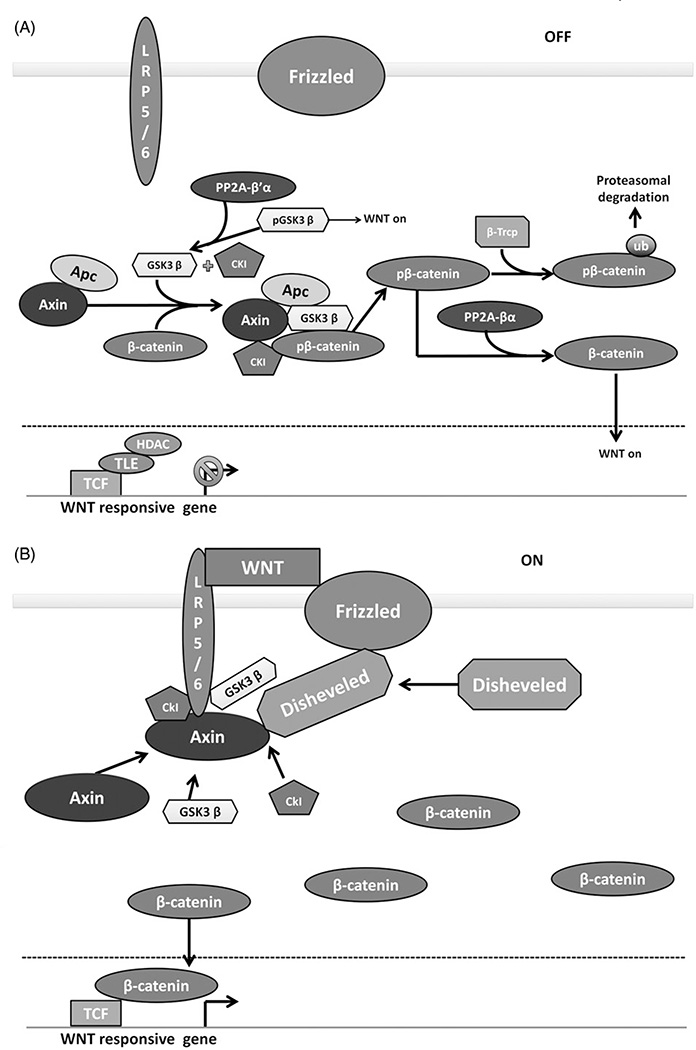
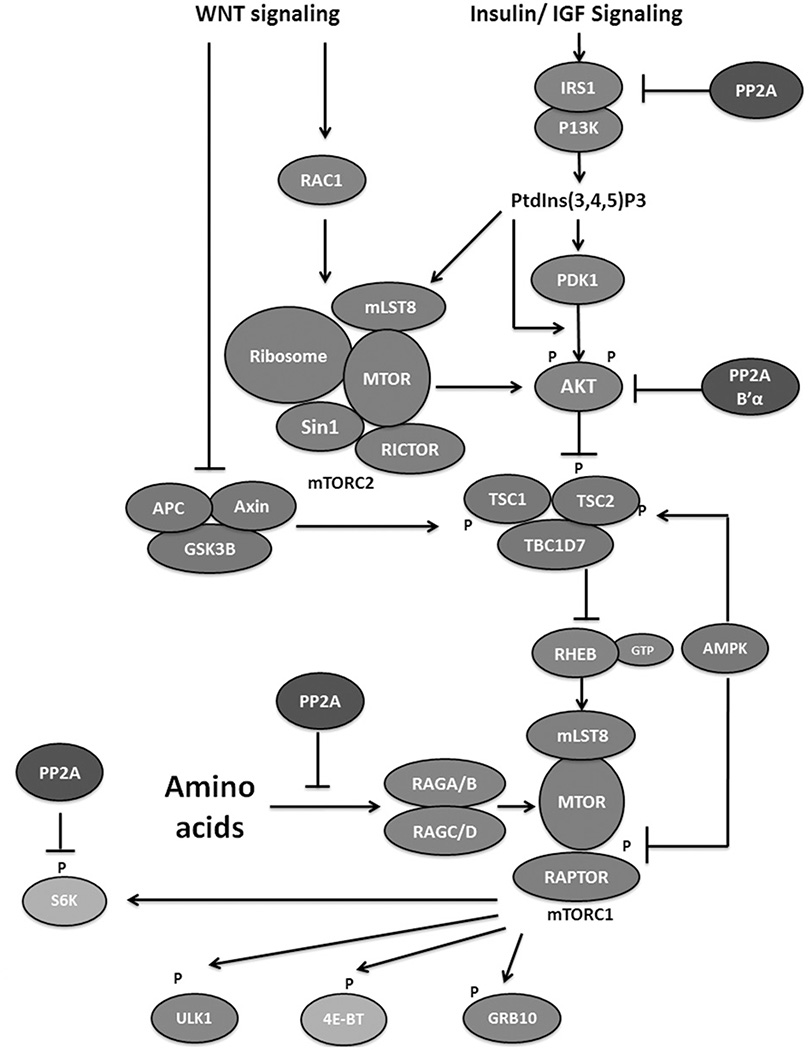
References
-
- Adams DG, Coffee RL, Jr, Zhang H, et al. Positive regulation of Raf1-MEK1/2-ERK1/2 signaling by protein serine/threonine phosphatase 2A holoenzymes. J Biol Chem. 2005;280:42644–42654. - PubMed
-
- Adler HT, Nallaseth FS, Walter G, Tkachuk DC. HRX leukemic fusion proteins form a heterocomplex with the leukemia-associated protein SET and protein phosphatase 2A. J Biol Chem. 1997;272:28407–28414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous