

The mechanics of state-dependent neural correlations
- PMID: 26906505
- PMCID: PMC5477791
- DOI: 10.1038/nn.4242
The mechanics of state-dependent neural correlations
Abstract
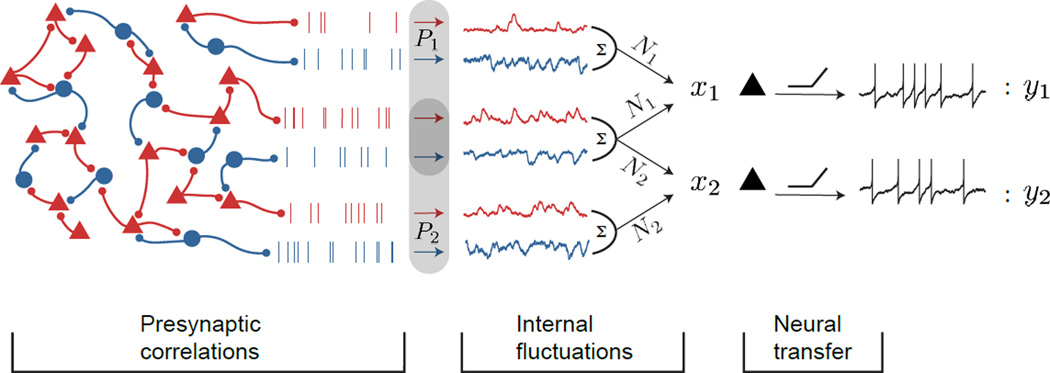
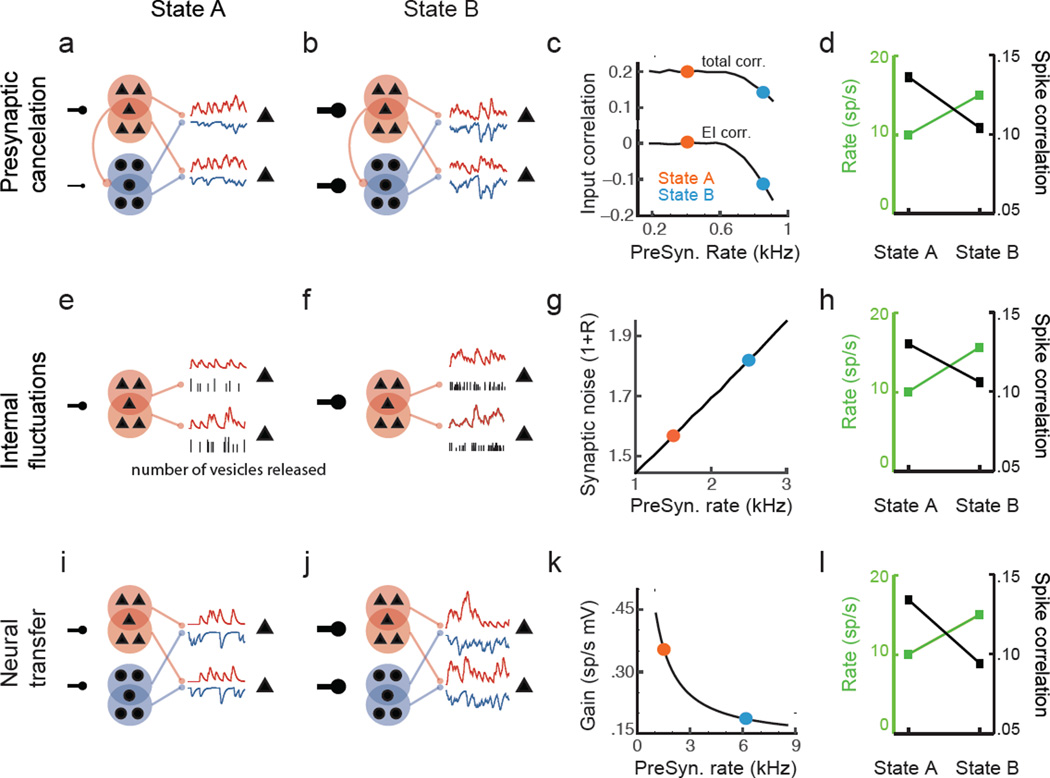
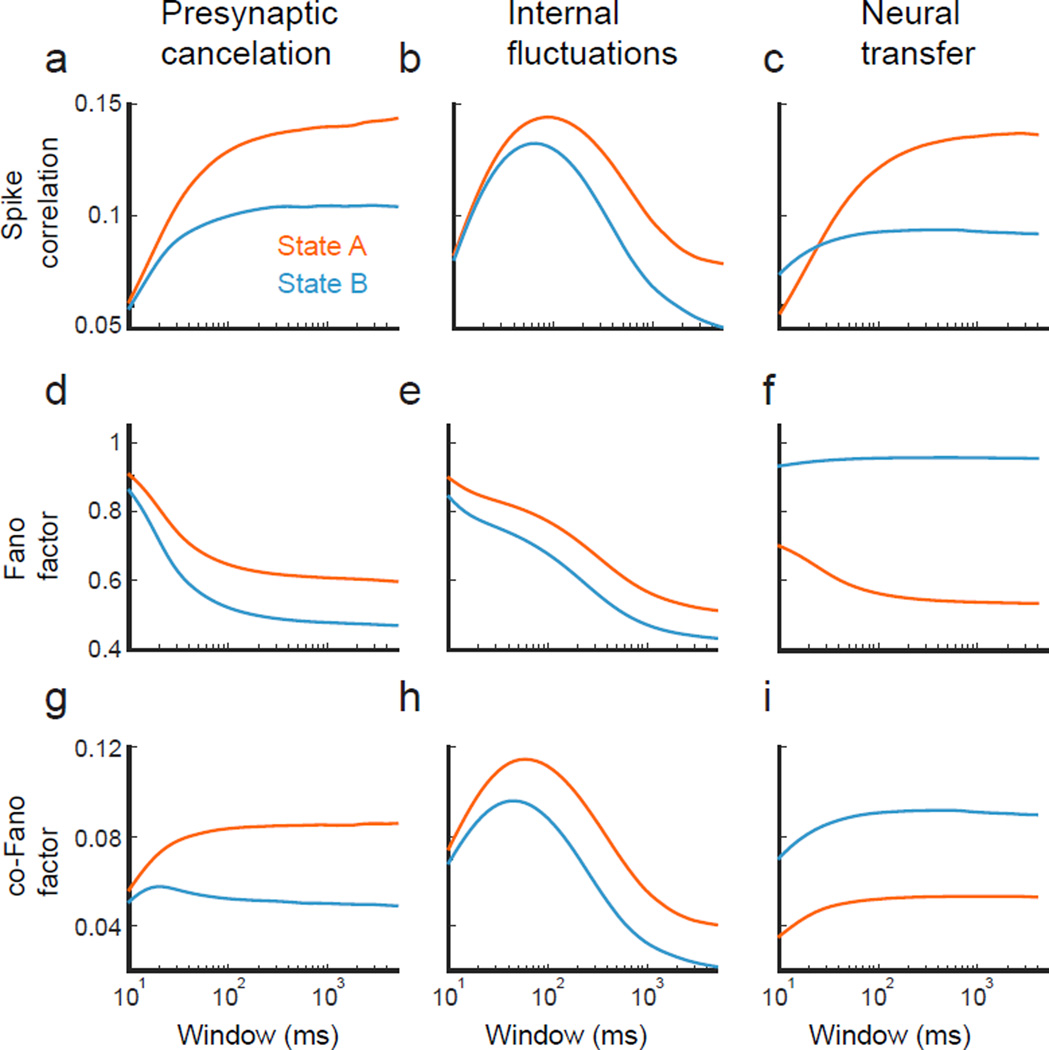
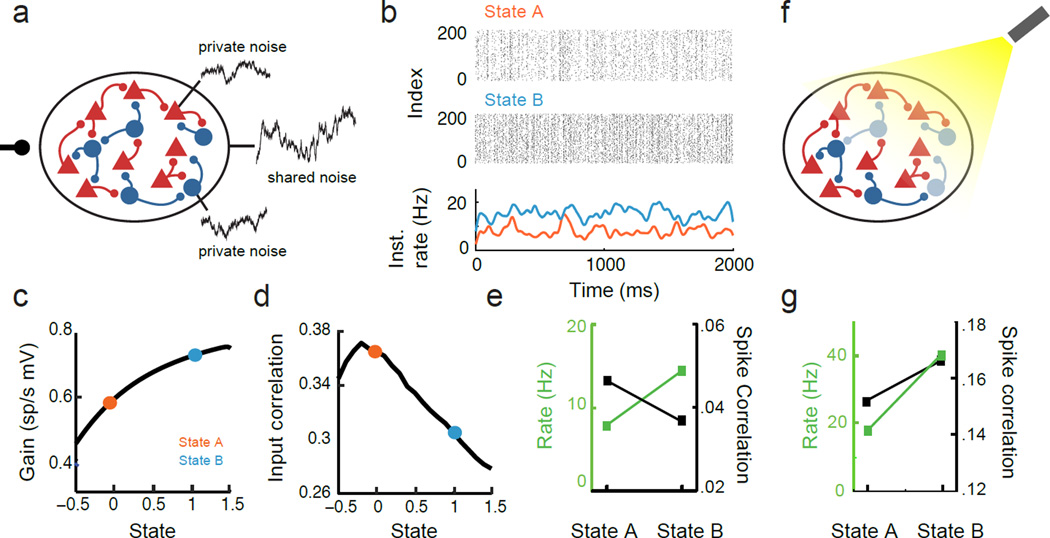
Simultaneous recordings from large neural populations are becoming increasingly common. An important feature of population activity is the trial-to-trial correlated fluctuation of spike train outputs from recorded neuron pairs. Similar to the firing rate of single neurons, correlated activity can be modulated by a number of factors, from changes in arousal and attentional state to learning and task engagement. However, the physiological mechanisms that underlie these changes are not fully understood. We review recent theoretical results that identify three separate mechanisms that modulate spike train correlations: changes in input correlations, internal fluctuations and the transfer function of single neurons. We first examine these mechanisms in feedforward pathways and then show how the same approach can explain the modulation of correlations in recurrent networks. Such mechanistic constraints on the modulation of population activity will be important in statistical analyses of high-dimensional neural data.
Figures
References
-
- Yuste R. From the neuron doctrine to neural networks. Nature Reviews Neuroscience. 2015;16:487–497. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
