

Does a Common Pathway Transduce Symbiotic Signals in Plant-Microbe Interactions?
- PMID: 26909085
- PMCID: PMC4754458
- DOI: 10.3389/fpls.2016.00096
Does a Common Pathway Transduce Symbiotic Signals in Plant-Microbe Interactions?
Abstract
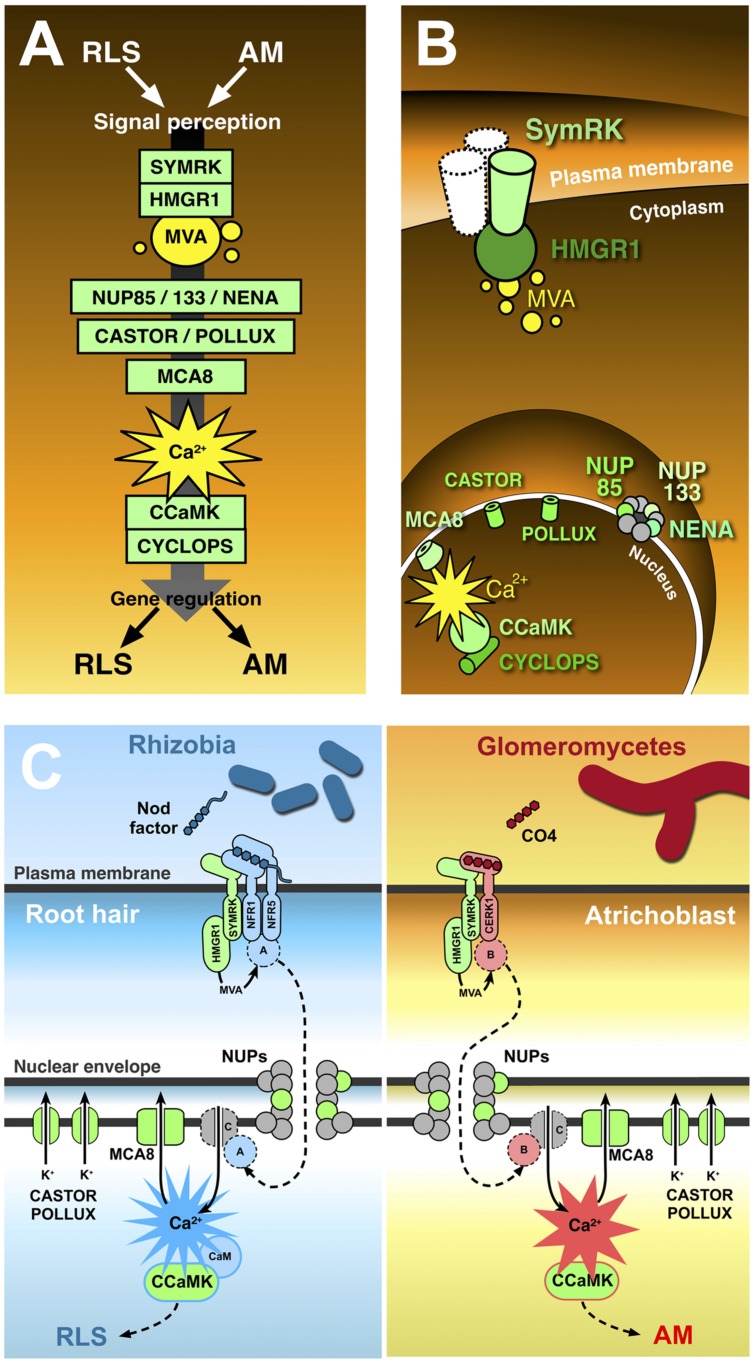
Recent years have witnessed major advances in our knowledge of plant mutualistic symbioses such as the rhizobium-legume symbiosis (RLS) and arbuscular mycorrhizas (AM). Some of these findings caused the revision of longstanding hypotheses, but one of the most solid theories is that a conserved set of plant proteins rules the transduction of symbiotic signals from beneficial glomeromycetes and rhizobia in a so-called common symbiotic pathway (CSP). Nevertheless, the picture still misses several elements, and a few crucial points remain unclear. How does one common pathway discriminate between - at least - two symbionts? Can we exclude that microbes other than AM fungi and rhizobia also use this pathway to communicate with their host plants? We here discuss the possibility that our current view is biased by a long-lasting focus on legumes, whose ability to develop both AM and RLS is an exception among plants and a recent innovation in their evolution; investigations in non-legumes are starting to place legume symbiotic signaling in a broader perspective. Furthermore, recent studies suggest that CSP proteins act in a wider scenario of symbiotic and non-symbiotic signaling. Overall, evidence is accumulating in favor of distinct activities for CSP proteins in AM and RLS, depending on the molecular and cellular context where they act.
Keywords: arbuscular mycorrhiza; legume nodulation; plant–microbe interactions; signaling pathways; symbiosis.
Figures
Similar articles
-
Lotus japonicus Symbiosis Genes Impact Microbial Interactions between Symbionts and Multikingdom Commensal Communities.mBio. 2019 Oct 8;10(5):e01833-19. doi: 10.1128/mBio.01833-19. mBio. 2019. PMID: 31594815 Free PMC article.
-
Signaling in the arbuscular mycorrhizal symbiosis.Annu Rev Microbiol. 2005;59:19-42. doi: 10.1146/annurev.micro.58.030603.123749. Annu Rev Microbiol. 2005. PMID: 16153162 Review.
-
The bifunctional plant receptor, OsCERK1, regulates both chitin-triggered immunity and arbuscular mycorrhizal symbiosis in rice.Plant Cell Physiol. 2014 Nov;55(11):1864-72. doi: 10.1093/pcp/pcu129. Epub 2014 Sep 17. Plant Cell Physiol. 2014. PMID: 25231970
-
[Comparative genetics and evolutionary morphology of symbiosis formed by plants with nitrogen-fixing microbes and endomycorrhizal fungi].Zh Obshch Biol. 2002 Nov-Dec;63(6):451-72. Zh Obshch Biol. 2002. PMID: 12510586 Review. Russian.
-
CERBERUS and NSP1 of Lotus japonicus are common symbiosis genes that modulate arbuscular mycorrhiza development.Plant Cell Physiol. 2013 Oct;54(10):1711-23. doi: 10.1093/pcp/pct114. Epub 2013 Aug 7. Plant Cell Physiol. 2013. PMID: 23926062
Cited by
-
Temporal change in chromatin accessibility predicts regulators of nodulation in Medicago truncatula.BMC Biol. 2022 Nov 9;20(1):252. doi: 10.1186/s12915-022-01450-9. BMC Biol. 2022. PMID: 36352404 Free PMC article.
-
Glycine max Homologs of DOESN'T MAKE INFECTIONS 1, 2, and 3 Function to Impair Heterodera glycines Parasitism While Also Regulating Mitogen Activated Protein Kinase Expression.Front Plant Sci. 2022 May 4;13:842597. doi: 10.3389/fpls.2022.842597. eCollection 2022. Front Plant Sci. 2022. PMID: 35599880 Free PMC article.
-
Transcription factors network in root endosymbiosis establishment and development.World J Microbiol Biotechnol. 2018 Feb 15;34(3):37. doi: 10.1007/s11274-018-2418-7. World J Microbiol Biotechnol. 2018. PMID: 29450655 Review.
-
Early Molecular Dialogue Between Legumes and Rhizobia: Why Are They So Important?Results Probl Cell Differ. 2020;69:409-419. doi: 10.1007/978-3-030-51849-3_15. Results Probl Cell Differ. 2020. PMID: 33263881
-
Determining the Site of Action of Strigolactones during Nodulation.Plant Physiol. 2017 Sep;175(1):529-542. doi: 10.1104/pp.17.00741. Epub 2017 Jul 27. Plant Physiol. 2017. PMID: 28751316 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources