

Structures of Two Melanoma-Associated Antigens Suggest Allosteric Regulation of Effector Binding
- PMID: 26910052
- PMCID: PMC4766014
- DOI: 10.1371/journal.pone.0148762
Structures of Two Melanoma-Associated Antigens Suggest Allosteric Regulation of Effector Binding
Abstract
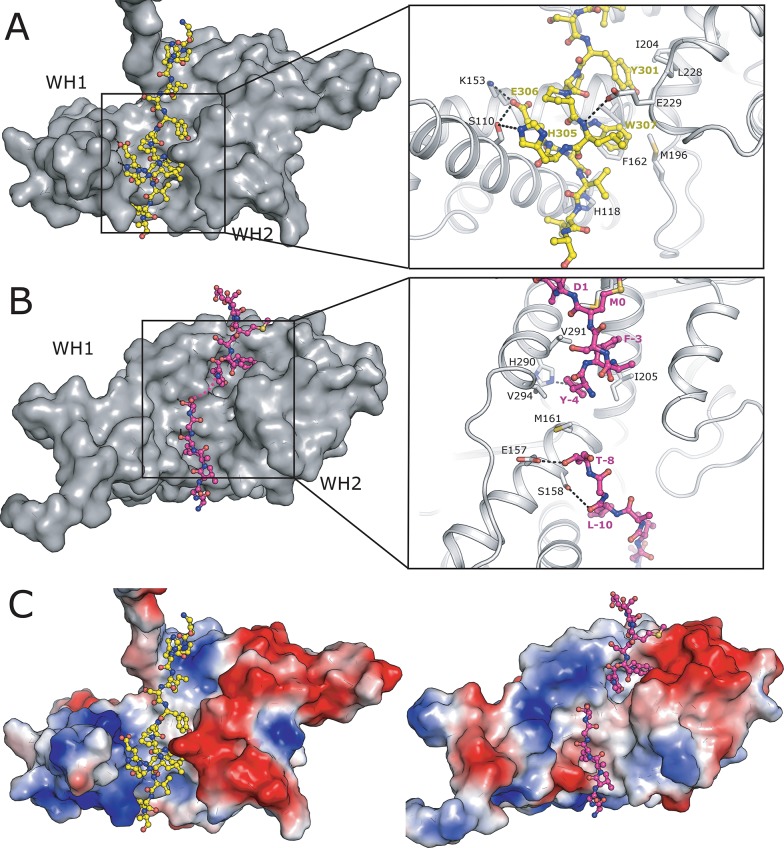
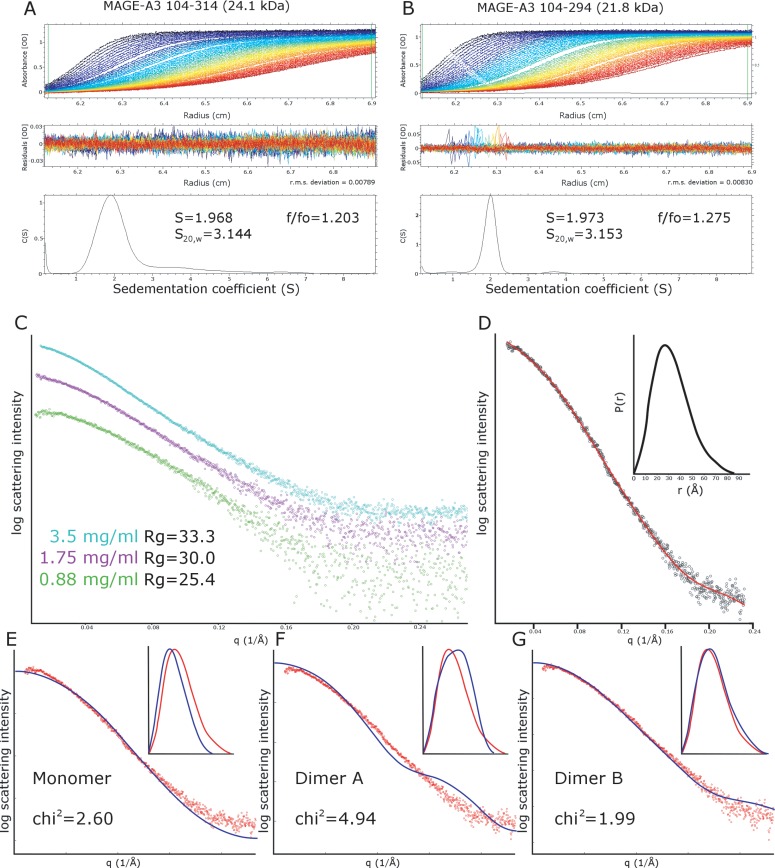
The MAGE (melanoma associated antigen) protein family are tumour-associated proteins normally present only in reproductive tissues such as germ cells of the testis. The human genome encodes over 60 MAGE genes of which one class (containing MAGE-A3 and MAGE-A4) are exclusively expressed in tumours, making them an attractive target for the development of targeted and immunotherapeutic cancer treatments. Some MAGE proteins are thought to play an active role in driving cancer, modulating the activity of E3 ubiquitin ligases on targets related to apoptosis. Here we determined the crystal structures of MAGE-A3 and MAGE-A4. Both proteins crystallized with a terminal peptide bound in a deep cleft between two tandem-arranged winged helix domains. MAGE-A3 (but not MAGE-A4), is predominantly dimeric in solution. Comparison of MAGE-A3 and MAGE-A3 with a structure of an effector-bound MAGE-G1 suggests that a major conformational rearrangement is required for binding, and that this conformational plasticity may be targeted by allosteric binders.
Conflict of interest statement
Figures







References
-
- van der Bruggen P, Traversari C, Chomez P, Lurquin C, De Plaen E, Van den Eynde B, et al. A gene encoding an antigen recognized by cytolytic T lymphocytes on a human melanoma. Science. 1991;254(5038):1643–7. . - PubMed
-
- Barker PA, Salehi A. The MAGE proteins: emerging roles in cell cycle progression, apoptosis, and neurogenetic disease. Journal of neuroscience research. 2002;67(6):705–12. . - PubMed
-
- Chomez P, De Backer O, Bertrand M, De Plaen E, Boon T, Lucas S. An overview of the MAGE gene family with the identification of all human members of the family. Cancer research. 2001;61(14):5544–51. . - PubMed
-
- De Smet C, Courtois SJ, Faraoni I, Lurquin C, Szikora JP, De Backer O, et al. Involvement of two Ets binding sites in the transcriptional activation of the MAGE1 gene. Immunogenetics. 1995;42(4):282–90. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
