

FOXC1 maintains the hair follicle stem cell niche and governs stem cell quiescence to preserve long-term tissue-regenerating potential
- PMID: 26912458
- PMCID: PMC4801248
- DOI: 10.1073/pnas.1601569113
FOXC1 maintains the hair follicle stem cell niche and governs stem cell quiescence to preserve long-term tissue-regenerating potential
Abstract
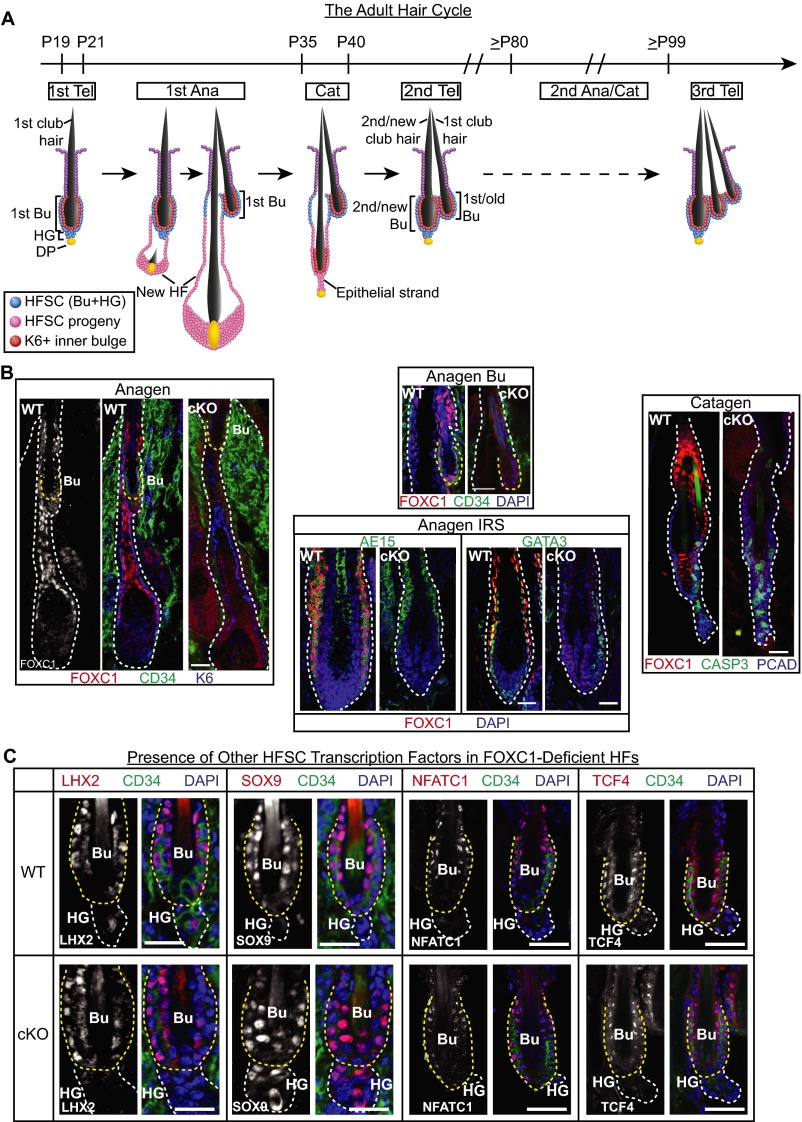
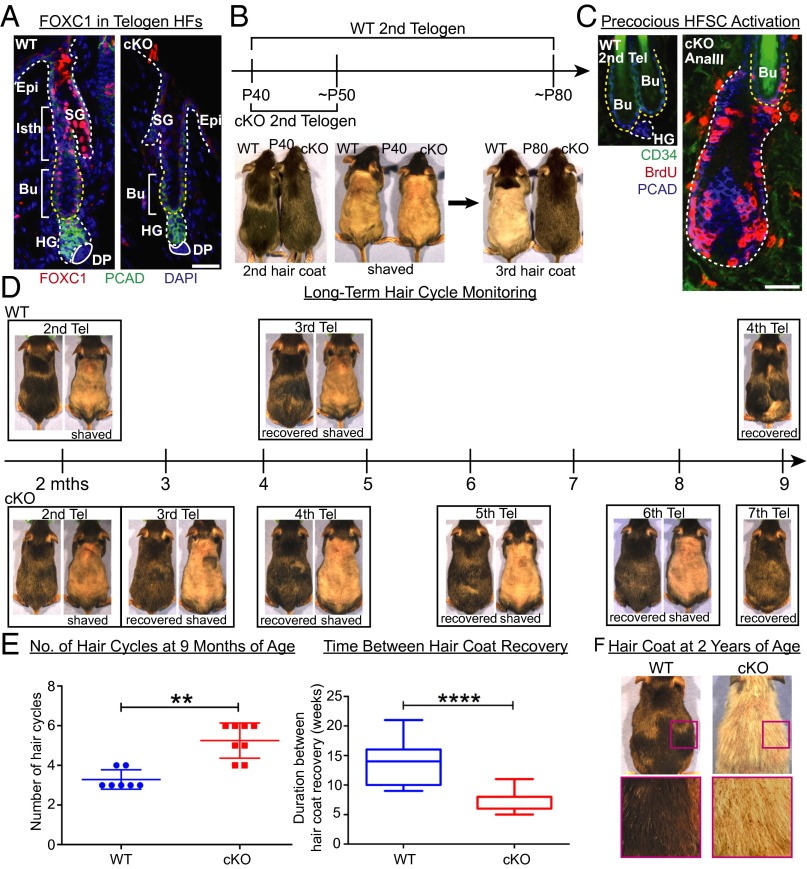
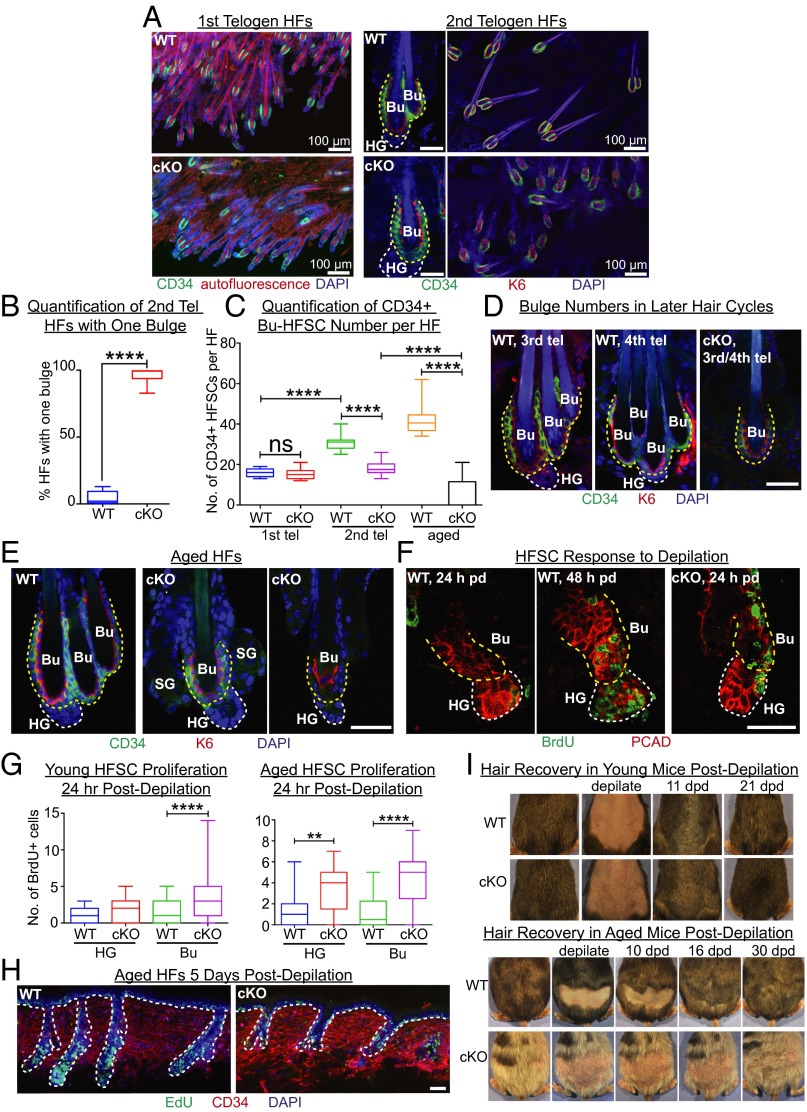
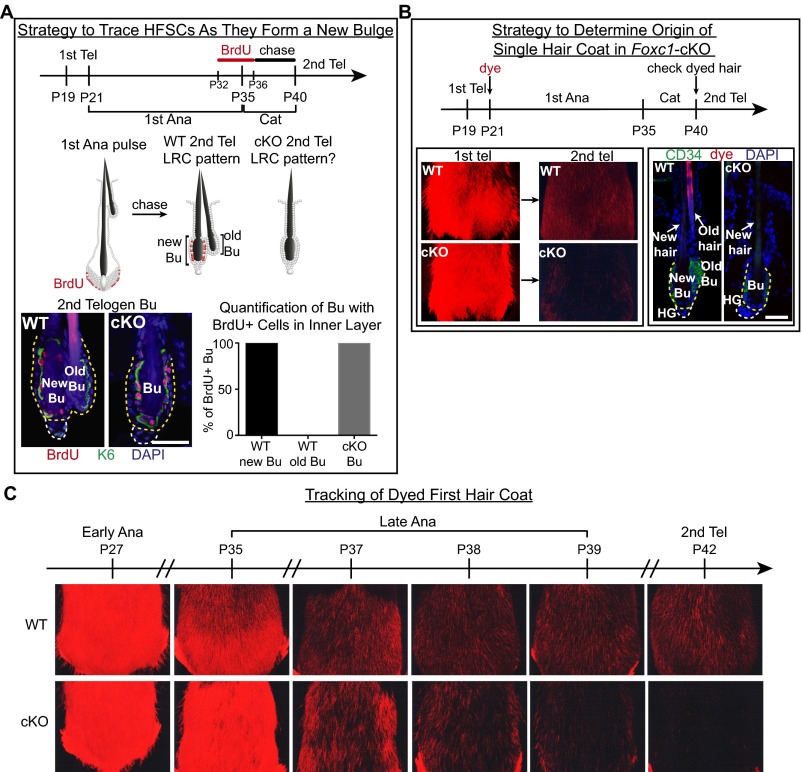
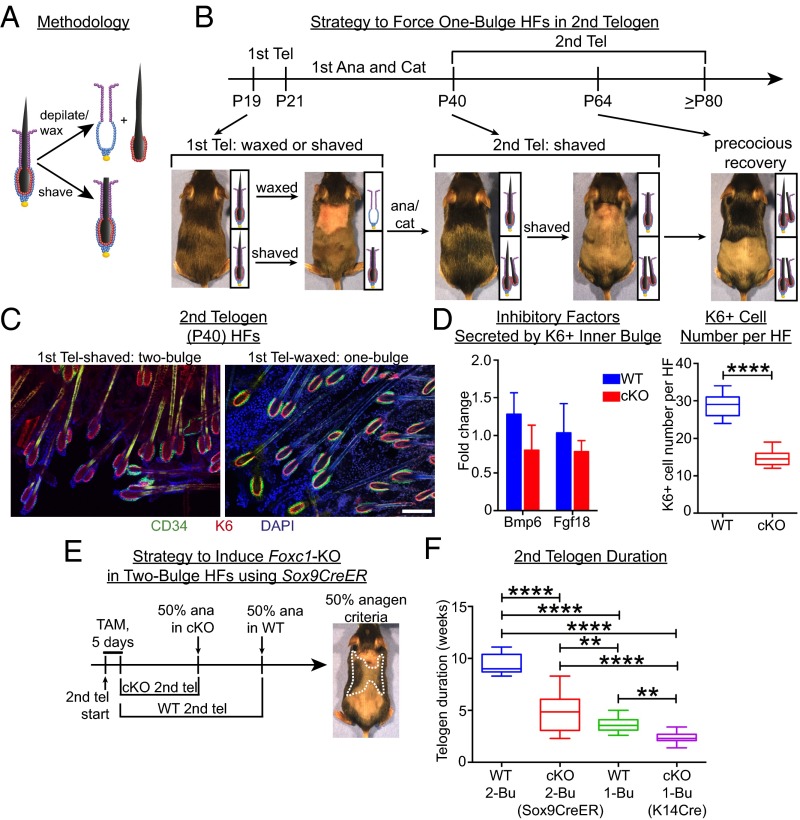
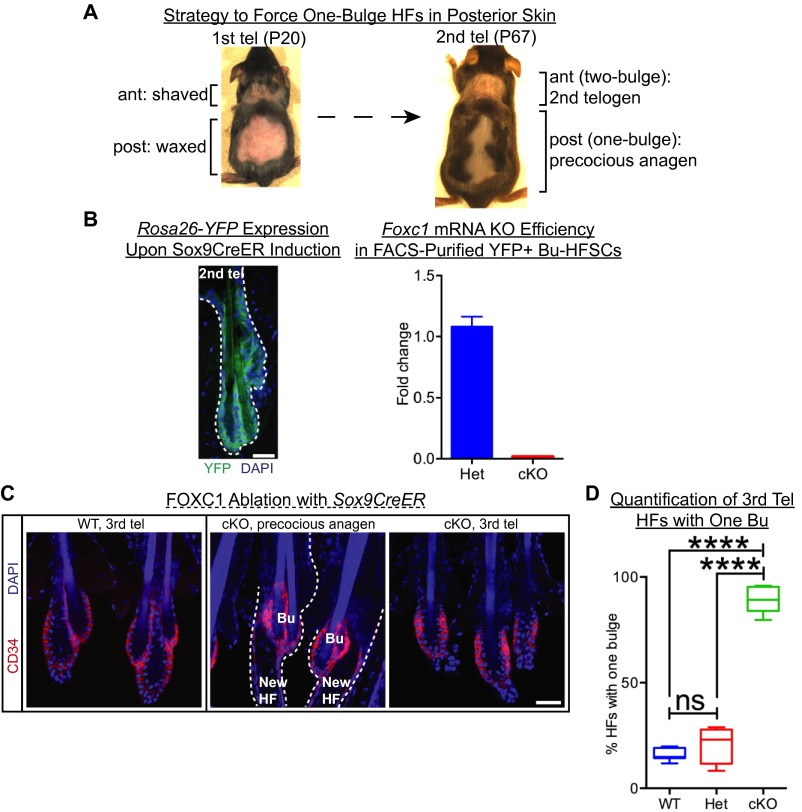
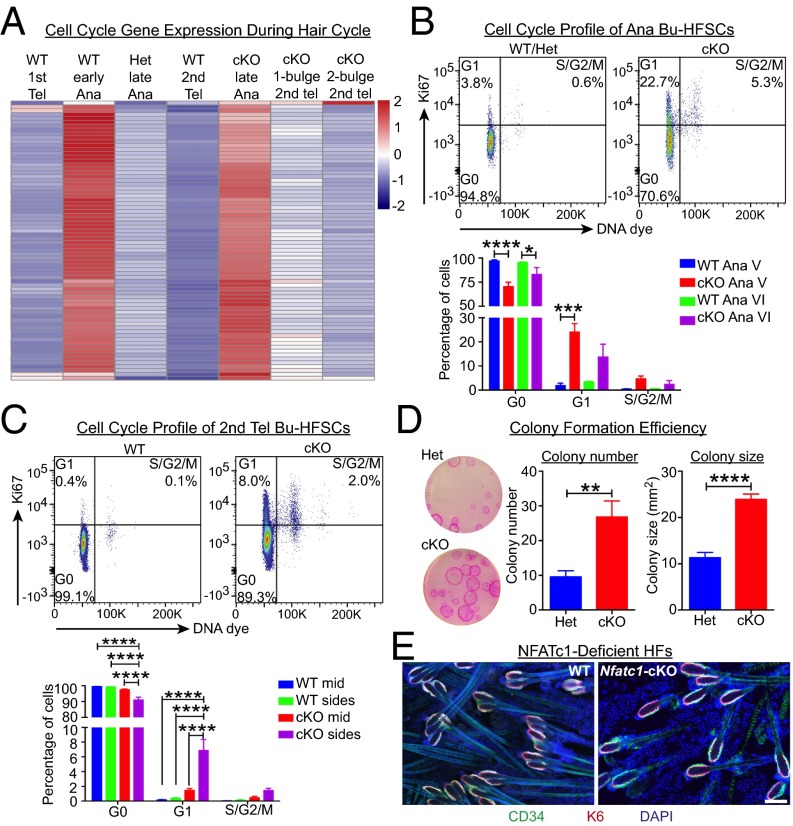
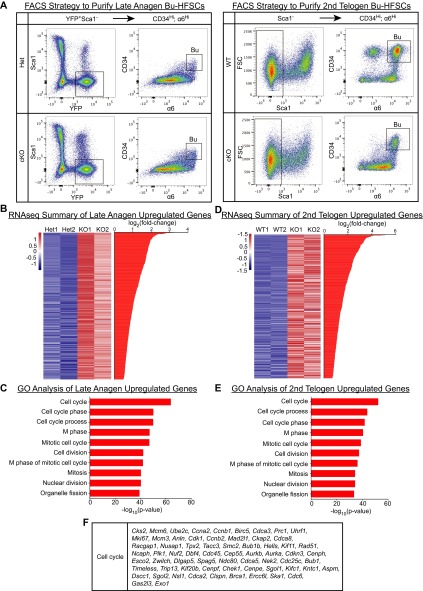
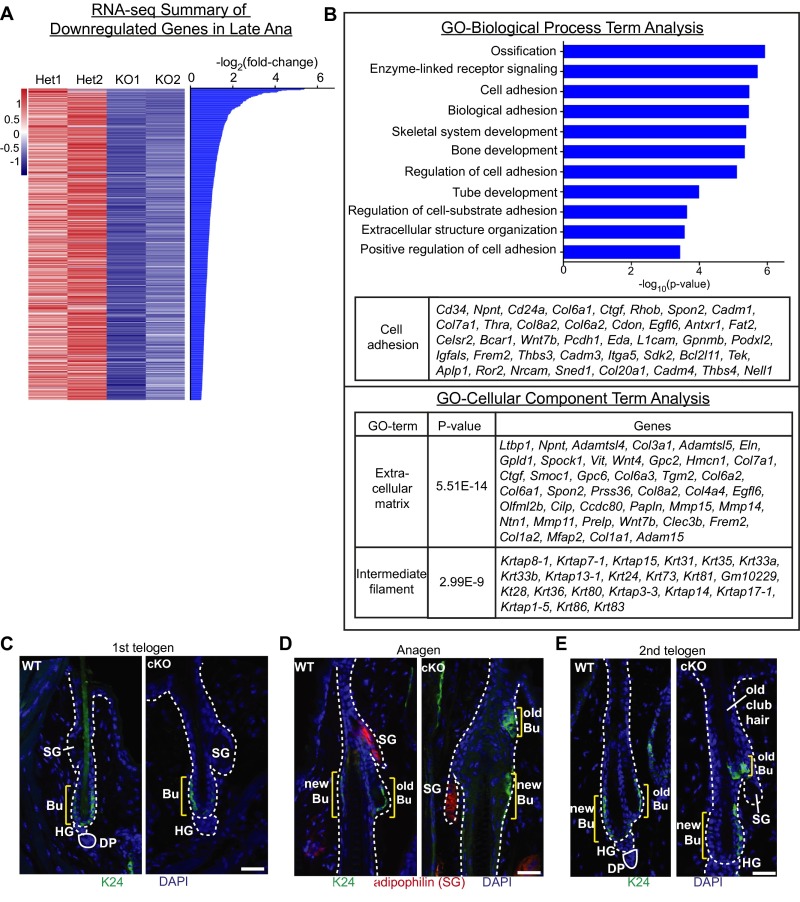
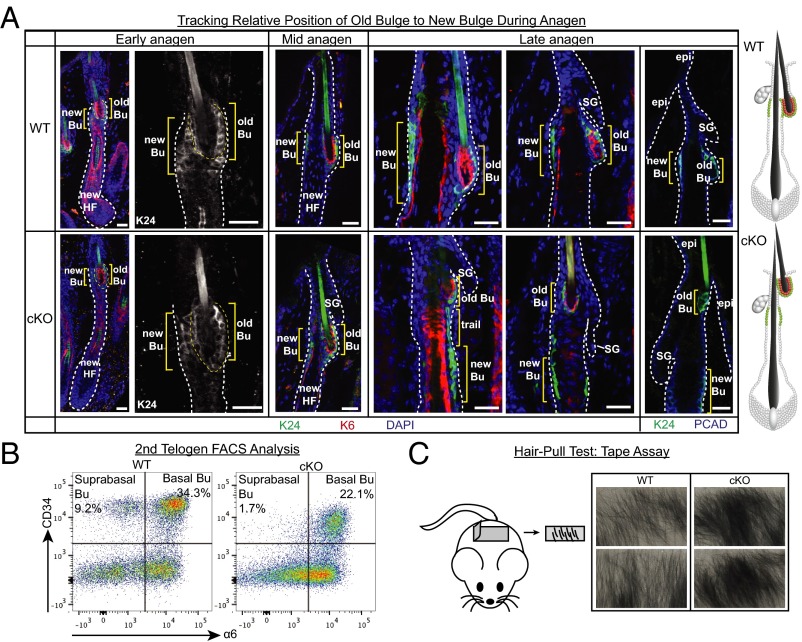
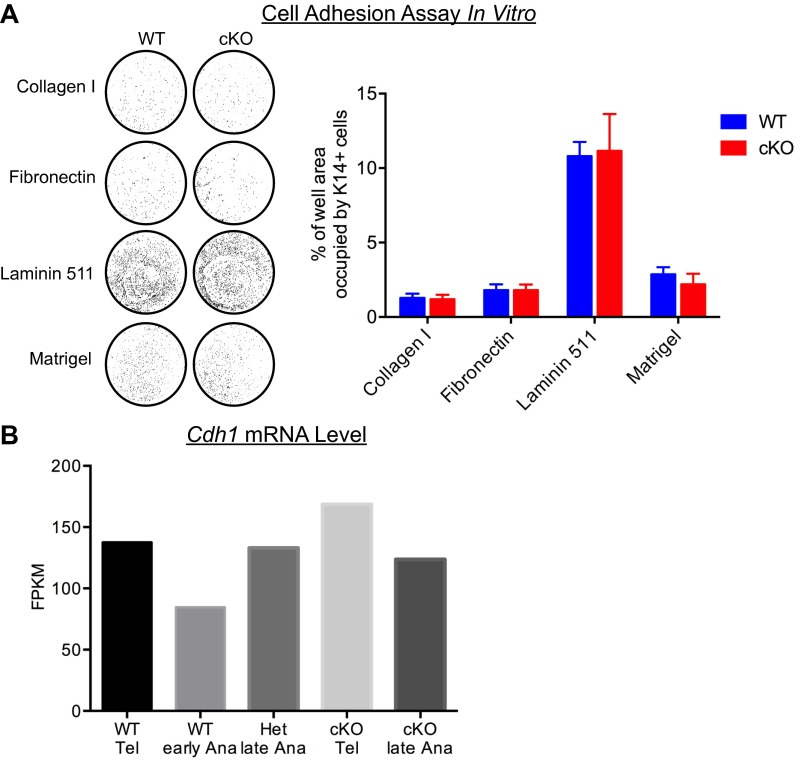
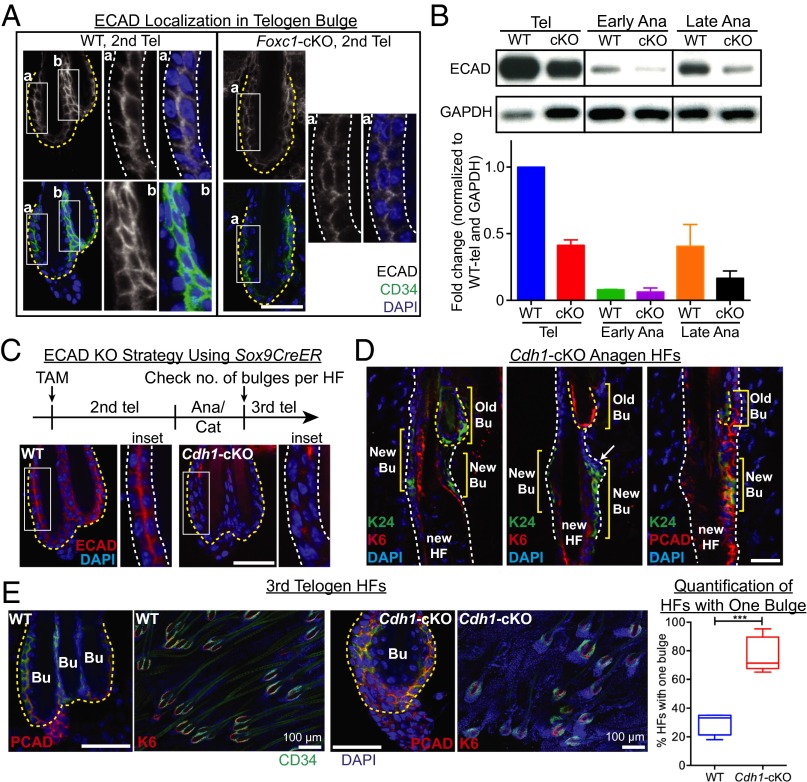
Adult tissue stem cells (SCs) reside in niches, which orchestrate SC behavior. SCs are typically used sparingly and exist in quiescence unless activated for tissue growth. Whether parsimonious SC use is essential to conserve long-term tissue-regenerating potential during normal homeostasis remains poorly understood. Here, we examine this issue by conditionally ablating a key transcription factor Forkhead box C1 (FOXC1) expressed in hair follicle SCs (HFSCs). FOXC1-deficient HFSCs spend less time in quiescence, leading to markedly shortened resting periods between hair cycles. The enhanced hair cycling accelerates HFSC expenditure, and impacts hair regeneration in aging mice. Interestingly, although FOXC1-deficient HFs can still form a new bulge that houses HFSCs for the next hair cycle, the older bulge is left unanchored. As the new hair emerges, the entire old bulge, including its reserve HFSCs and SC-inhibitory inner cell layer, is lost. We trace this mechanism first, to a marked increase in cell cycle-associated transcripts upon Foxc1 ablation, and second, to a downstream reduction in E-cadherin-mediated inter-SC adhesion. Finally, we show that when the old bulge is lost with each hair cycle, overall levels of SC-inhibitory factors are reduced, further lowering the threshold for HFSC activity. Taken together, our findings suggest that HFSCs have restricted potential in vivo, which they conserve by coupling quiescence to adhesion-mediated niche maintenance, thereby achieving long-term tissue homeostasis.
Keywords: FOXC1; aging; hair follicle; quiescence; stem cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Orford KW, Scadden DT. Deconstructing stem cell self-renewal: Genetic insights into cell-cycle regulation. Nat Rev Genet. 2008;9(2):115–128. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
