

Glycolysis and the pentose phosphate pathway are differentially associated with the dichotomous regulation of glioblastoma cell migration versus proliferation
- PMID: 26917237
- PMCID: PMC4998991
- DOI: 10.1093/neuonc/now024
Glycolysis and the pentose phosphate pathway are differentially associated with the dichotomous regulation of glioblastoma cell migration versus proliferation
Abstract
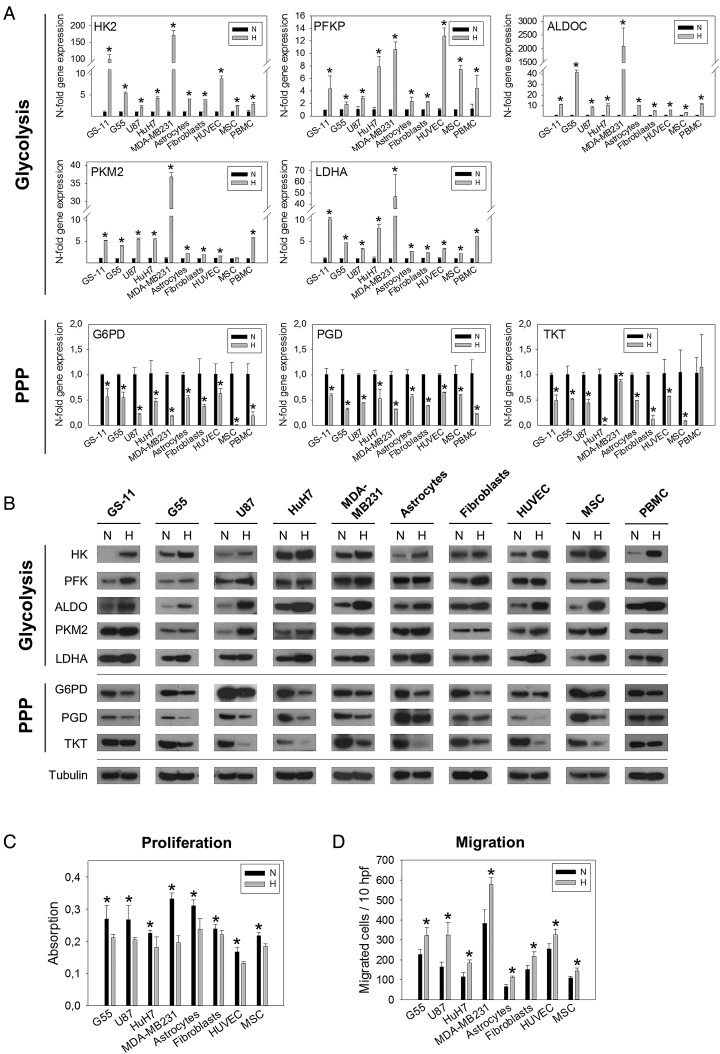
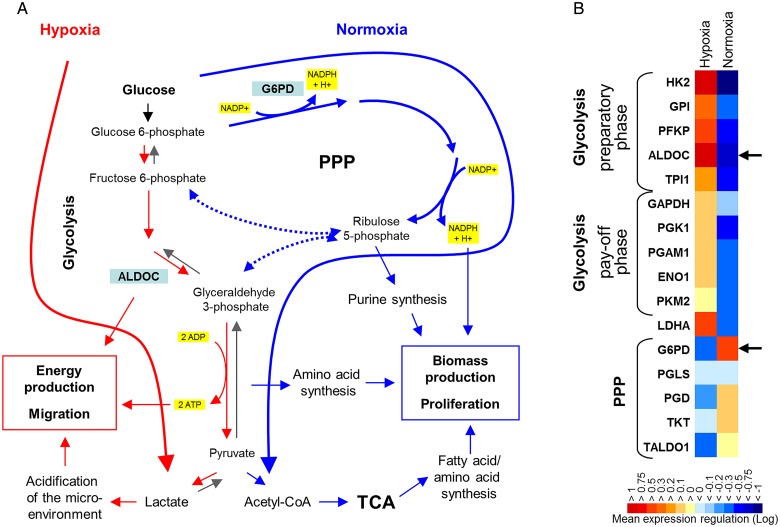
Background: The dichotomy between glioblastoma cell migration and proliferation is regulated by various parameters including oxygen tension. In glioblastoma stem-like cells, hypoxia induces downregulation of pentose phosphate pathway (PPP) enzymes and a flux shift towards glycolysis. We investigated whether the 2 parallel glucose metabolic pathways are intrinsically linked with cell function and whether these pathways are mechanistically involved in regulating functional programs.
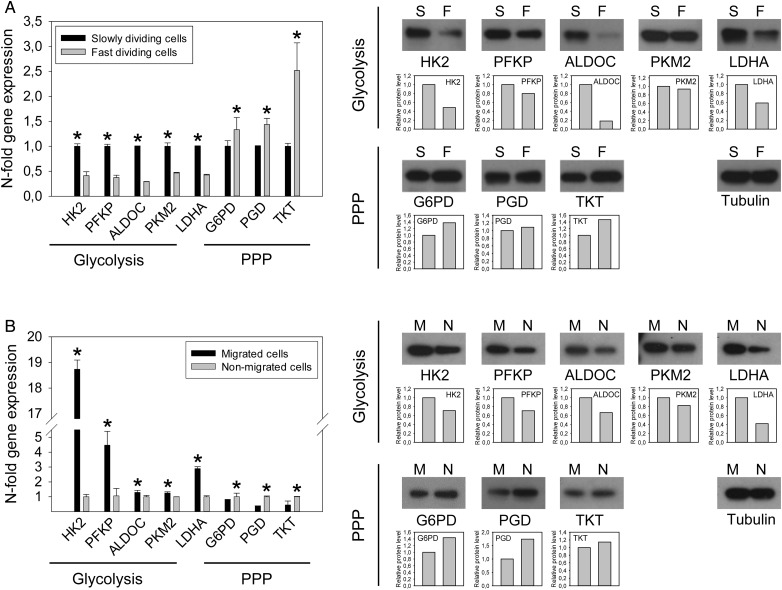
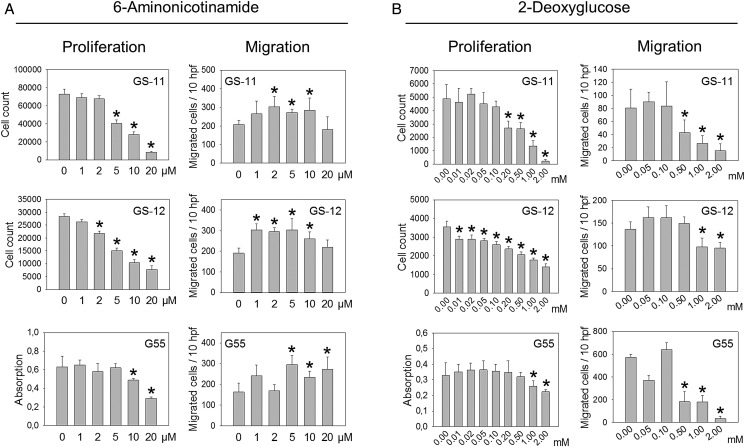
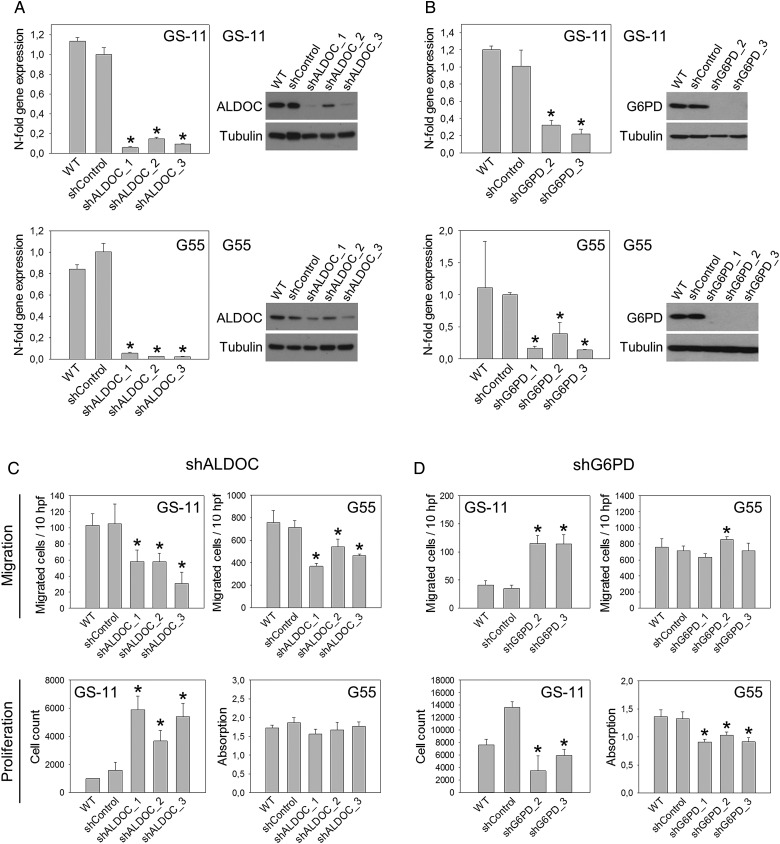
Methods: Enzyme expression, migration, and proliferation under hypoxia were studied in multiple cell types. Rapidly and slowly dividing or migrating glioblastoma cells were separated, and enzyme profiles were compared. Glucose-6-phosphate dehydrogenase (G6PD) and Aldolase C (ALDOC), the most strongly inversely regulated PPP and glycolysis enzymes, were knocked down by short hairpin RNA.
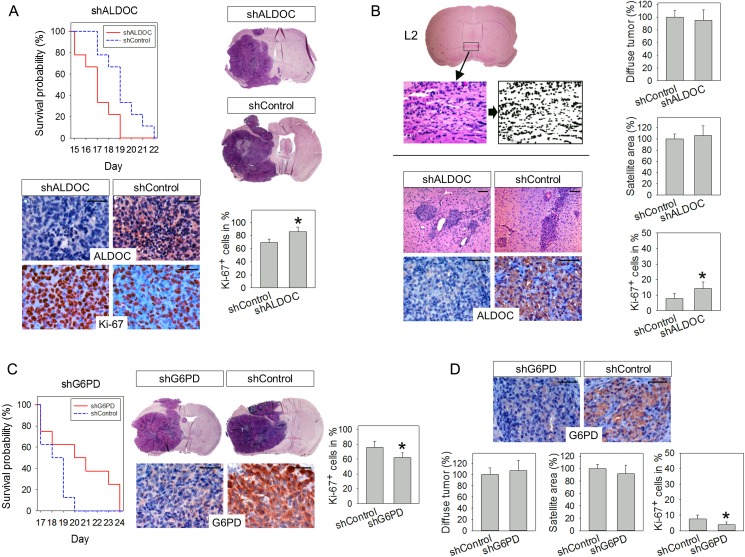
Results: Hypoxia caused downregulation of PPP enzymes and upregulation of glycolysis enzymes in a broad spectrum of cancer and nonneoplastic cells and consistently stimulated migration while reducing proliferation. PPP enzyme expression was increased in rapidly dividing glioblastoma cells, whereas glycolysis enzymes were decreased. Conversely, glycolysis enzymes were elevated in migrating cells, whereas PPP enzymes were diminished. Knockdown of G6PD reduced glioblastoma cell proliferation, whereas ALDOC knockdown decreased migration. Enzyme inhibitors had similar effects. G6PD knockdown in a highly proliferative but noninvasive glioblastoma cell line resulted in prolonged survival of mice with intracerebral xenografts, whereas ALDOC knockdown shortened survival. In a highly invasive glioblastoma xenograft model, tumor burden was unchanged by either knockdown.
Conclusions: Cell function and metabolic state are coupled independently of hypoxia, and glucose metabolic pathways are causatively involved in regulating "go or grow" cellular programs.
Keywords: glioma; glycolysis; invasion; metabolism; pentose phosphate pathway.
© The Author(s) 2016. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Giese A, Bjerkvig R, Berens ME, Westphal M. Cost of migration: invasion of malignant gliomas and implications for treatment. J Clin Oncol. 2003;21 (8):1624–1636. - PubMed
-
- Kunkel P, Ulbricht U, Bohlen P et al. . Inhibition of glioma angiogenesis and growth in vivo by systemic treatment with a monoclonal antibody against vascular endothelial growth factor receptor-2. Cancer Res. 2001;61 (18):6624–6628. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
