

Diverse roles for ionotropic glutamate receptors on inhibitory interneurons in developing and adult brain
- PMID: 26918438
- PMCID: PMC5043048
- DOI: 10.1113/JP271764
Diverse roles for ionotropic glutamate receptors on inhibitory interneurons in developing and adult brain
Abstract
Glutamate receptor-mediated recruitment of GABAergic inhibitory interneurons is a critical determinant of network processing. Early studies observed that many, but not all, interneuron glutamatergic synapses contain AMPA receptors that are GluA2-subunit lacking and Ca(2+) permeable, making them distinct from AMPA receptors at most principal cell synapses. Subsequent studies demonstrated considerable alignment of synaptic AMPA and NMDA receptor subunit composition within specific subtypes of interneurons, suggesting that both receptor expression profiles are developmentally and functionally linked. Indeed glutamate receptor expression profiles are largely predicted by the embryonic origins of cortical interneurons within the medial and caudal ganglionic eminences of the developing telencephalon. Distinct complements of AMPA and NMDA receptors within different interneuron subpopulations contribute to the differential recruitment of functionally divergent interneuron subtypes by common afferent inputs for appropriate feed-forward and feedback inhibitory drive and network entrainment. In contrast, the lesser-studied kainate receptors, which are often present at both pre- and postsynaptic sites, appear to follow an independent developmental expression profile. Loss of specific ionotropic glutamate receptor (iGluR) subunits during interneuron development has dramatic consequences for both cellular and network function, often precipitating circuit inhibition-excitation imbalances and in some cases lethality. Here we briefly review recent findings highlighting the roles of iGluRs in interneuron development.
Published 2016. This article is a U.S. Government work and is in the public domain in the USA.
Figures
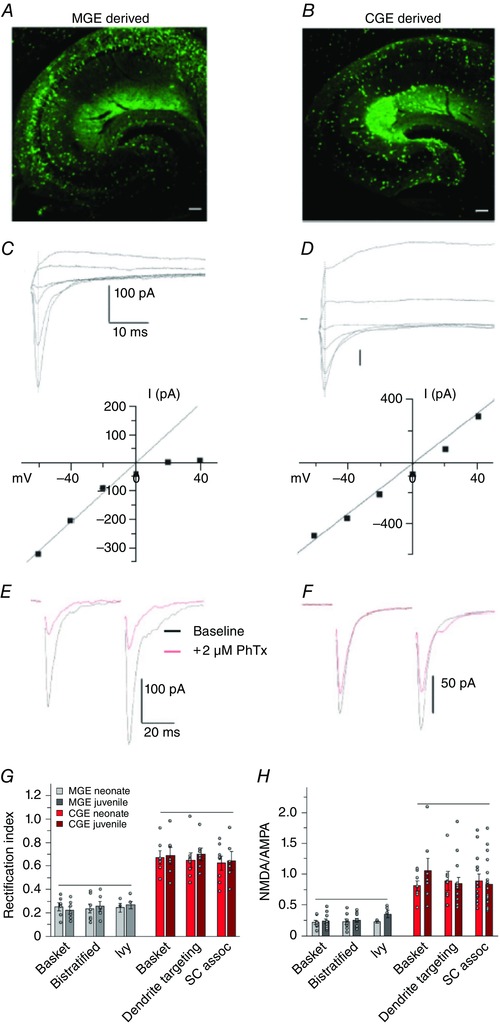
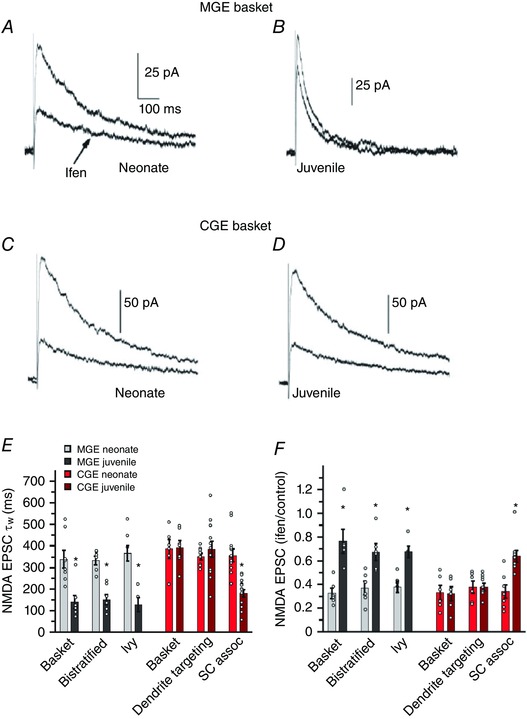
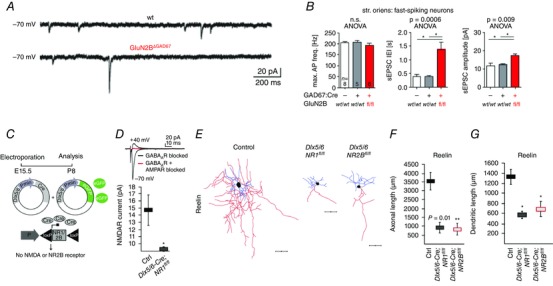
References
-
- Allen K & Monyer H (2015). Interneuron control of hippocampal oscillations. Curr Opin Neurobiol 31, 81–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous