

Liver fatty acid-binding protein (L-FABP) promotes cellular angiogenesis and migration in hepatocellular carcinoma
- PMID: 26919097
- PMCID: PMC4951284
- DOI: 10.18632/oncotarget.7571
Liver fatty acid-binding protein (L-FABP) promotes cellular angiogenesis and migration in hepatocellular carcinoma
Abstract
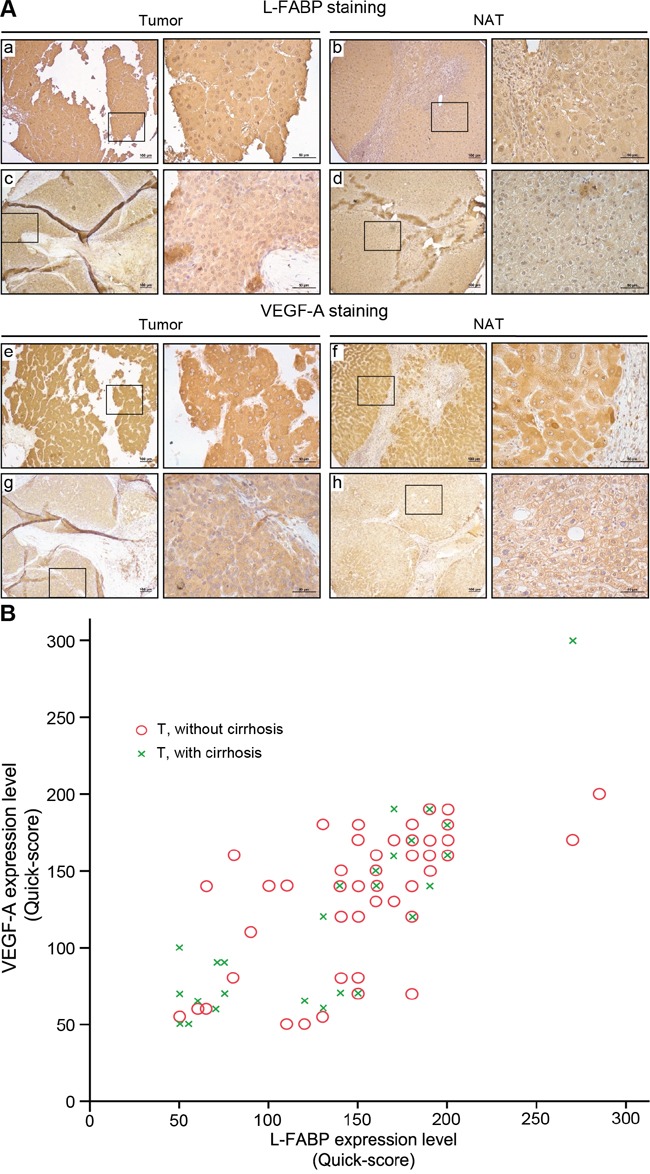
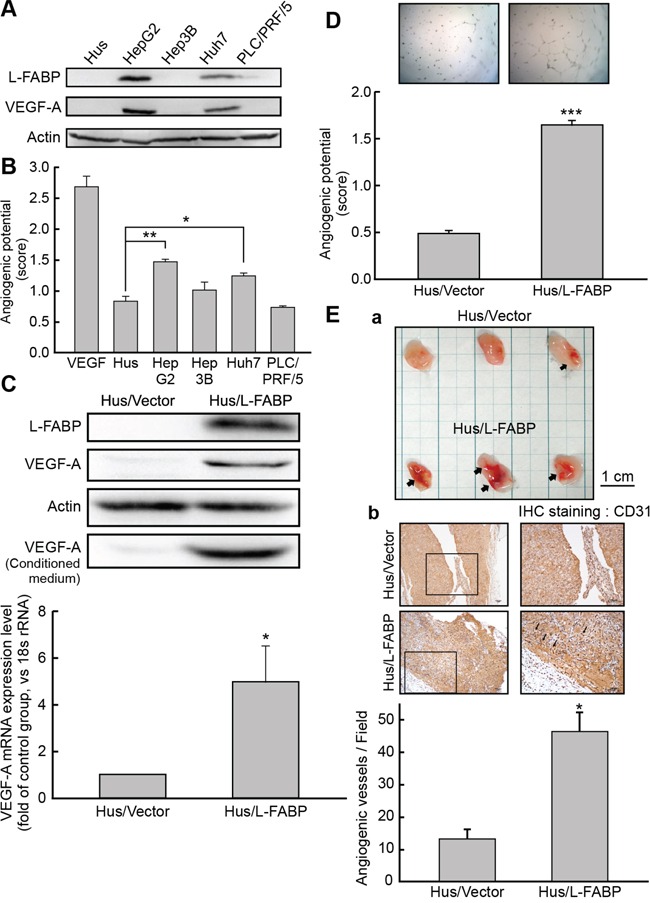
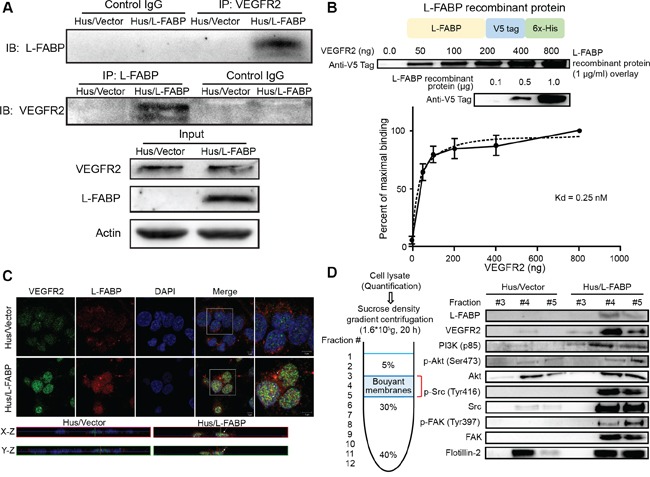
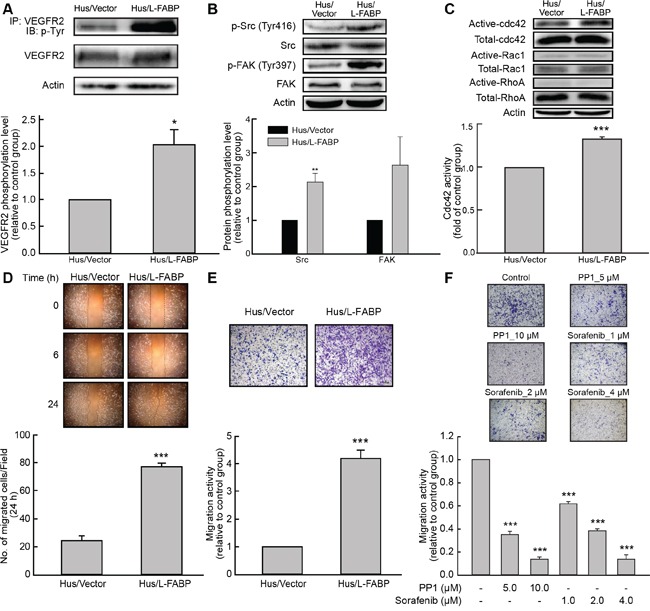
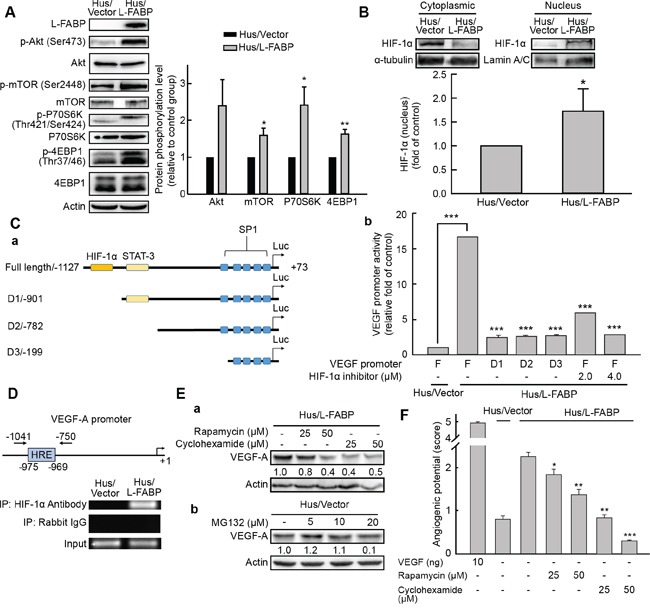
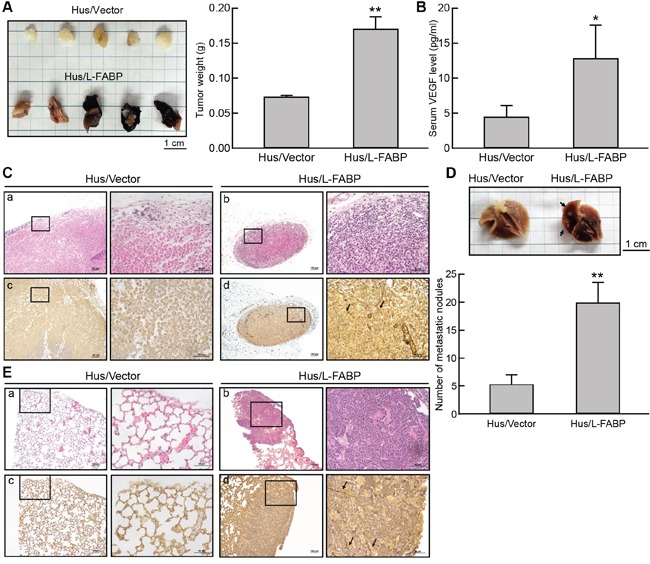
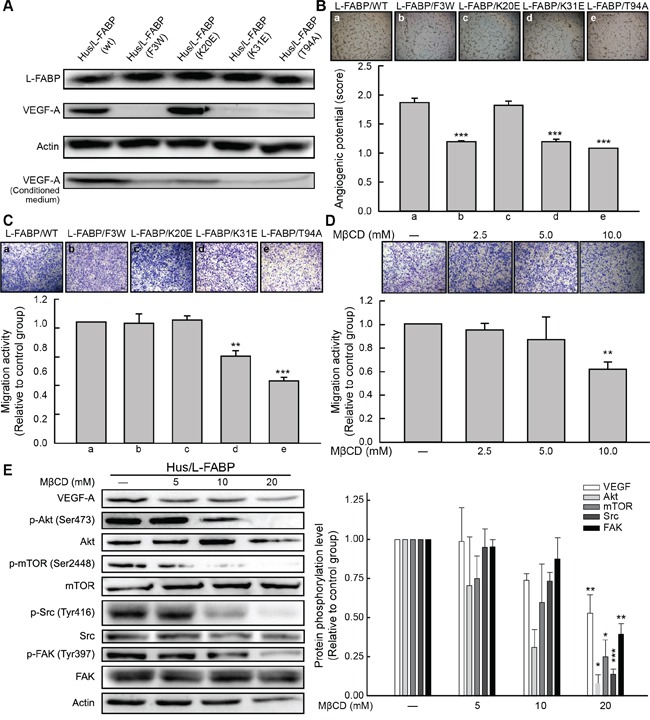
Liver fatty acid-binding protein (L-FABP) is abundant in hepatocytes and known to be involved in lipid metabolism. Overexpression of L-FABP has been reported in various cancers; however, its role in hepatocellular carcinoma (HCC) remains unclear. In this study, we investigated L-FABP and its association with vascular endothelial growth factors (VEGFs) in 90 HCC patients. We found that L-FABP was highly expressed in their HCC tissues, and that this expression was positively correlated with that of VEGF-A. Additionally, L-FABP significantly promoted tumor growth and metastasis in a xenograft mouse model. We also assessed the mechanisms of L-FABP activity in tumorigenesis; L-FABP was found to associate with VEGFR2 on membrane rafts and subsequently activate the Akt/mTOR/P70S6K/4EBP1 and Src/FAK/cdc42 pathways, which resulted in up-regulation of VEGF-A accompanied by an increase in both angiogenic potential and migration activity. Our results thus suggest that L-FABP could be a potential target for HCC chemotherapy.
Keywords: angiogenesis; hepatocellular carcinoma; liver fatty acid-binding protein; vascular endothelial growth factor.
Conflict of interest statement
No conflict of interest exists in the submission of this manuscript, and the manuscript was approved by all authors for publication.
Figures
References
-
- Kaseb AO, Hanbali A, Cotant M, Hassan MM, Wollner I, Philip PA. Vascular endothelial growth factor in the management of hepatocellular carcinoma: a review of literature. Cancer. 2009;115:4895–4906. - PubMed
-
- Romanque P, Piguet AC, Dufour JF. Targeting vessels to treat hepatocellular carcinoma. Clin Sci (Lond) 2008;114:467–477. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
