

Morpho-Physiological and Proteome Level Responses to Cadmium Stress in Sorghum
- PMID: 26919231
- PMCID: PMC4769174
- DOI: 10.1371/journal.pone.0150431
Morpho-Physiological and Proteome Level Responses to Cadmium Stress in Sorghum
Abstract
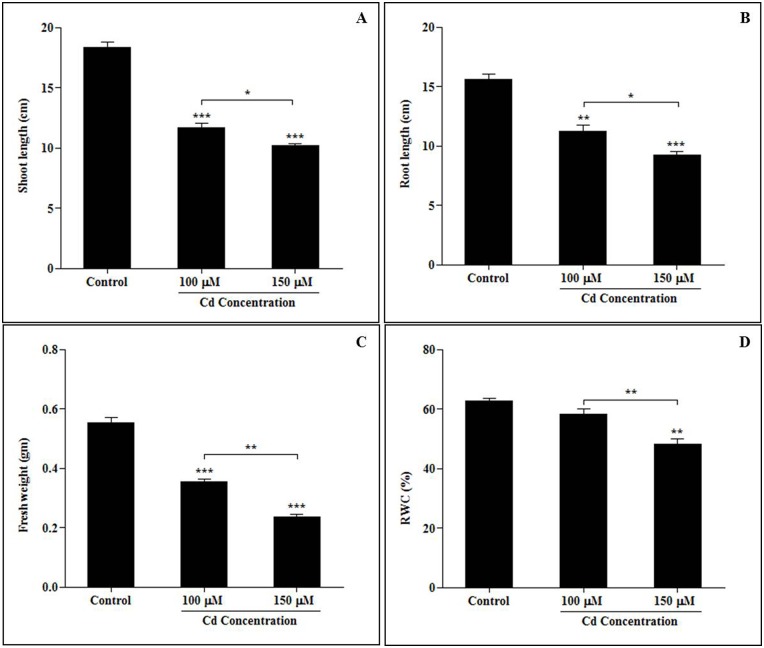
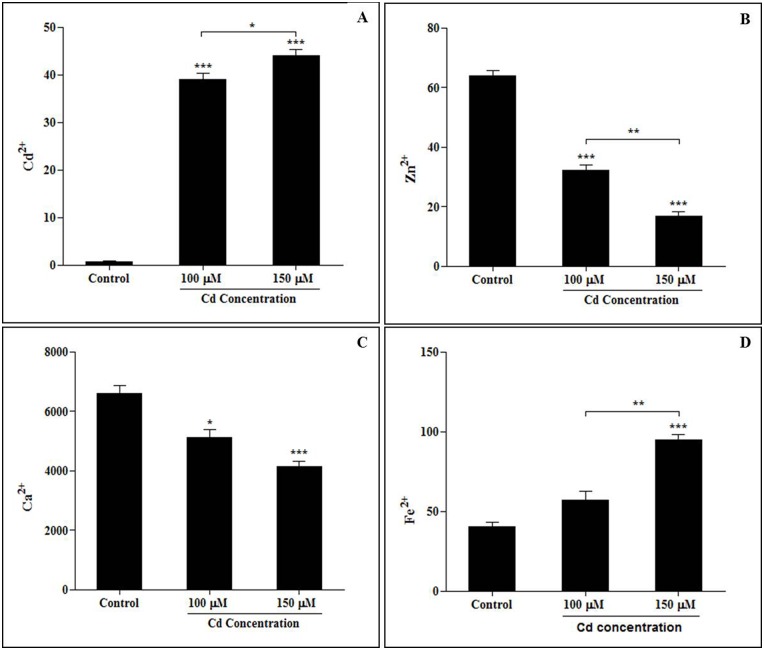
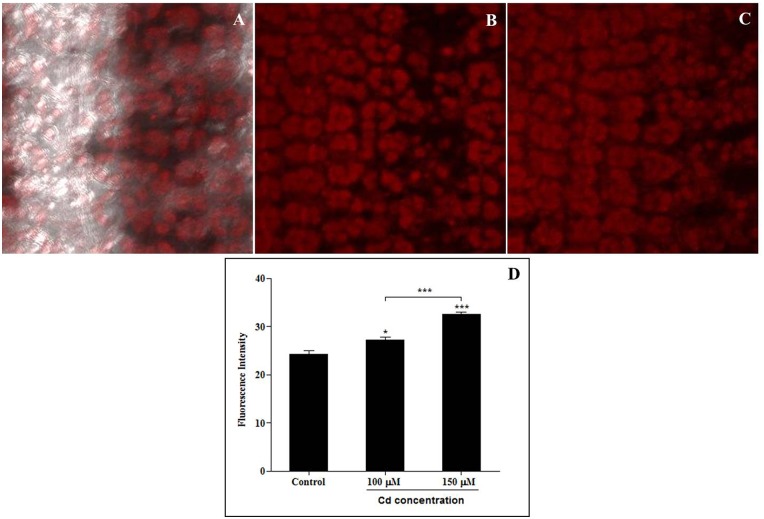
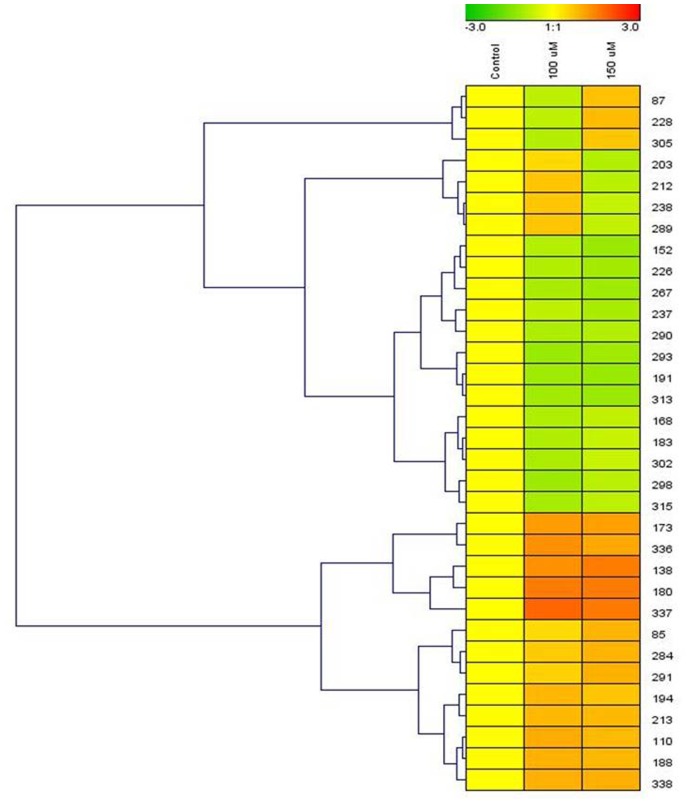
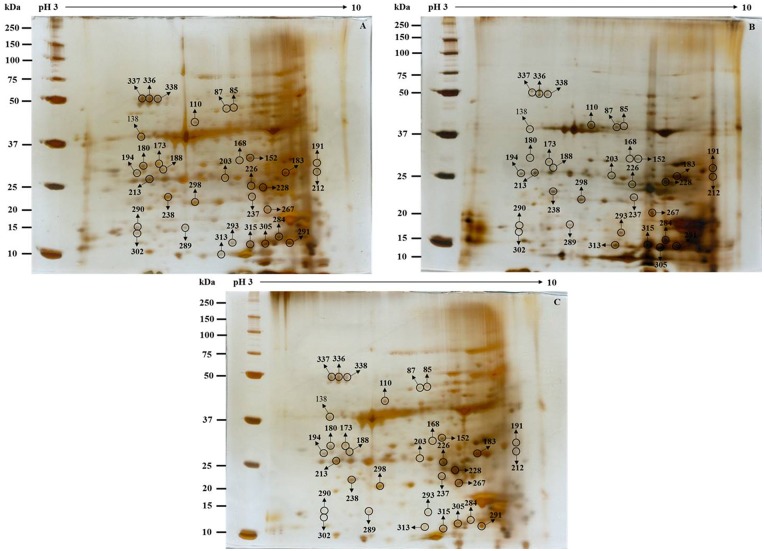
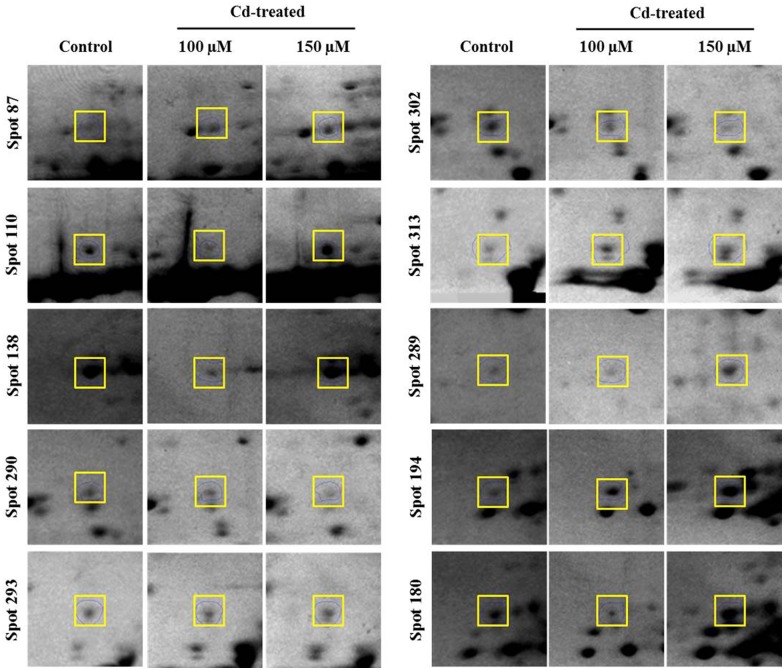
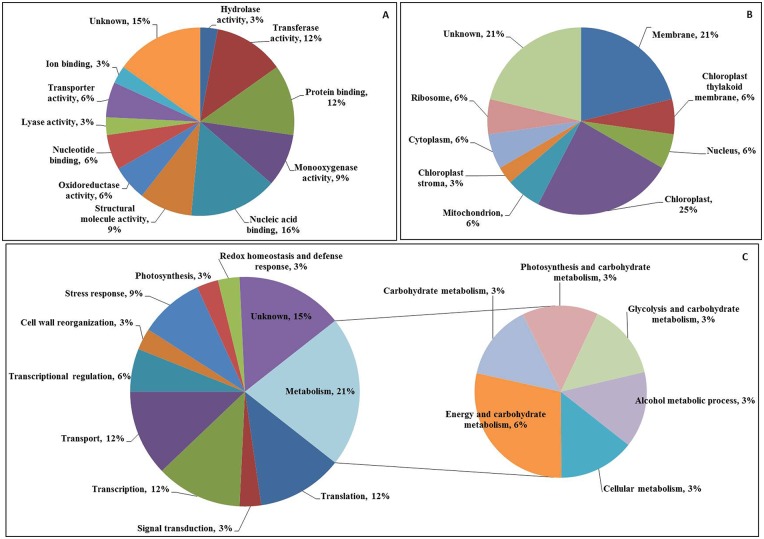
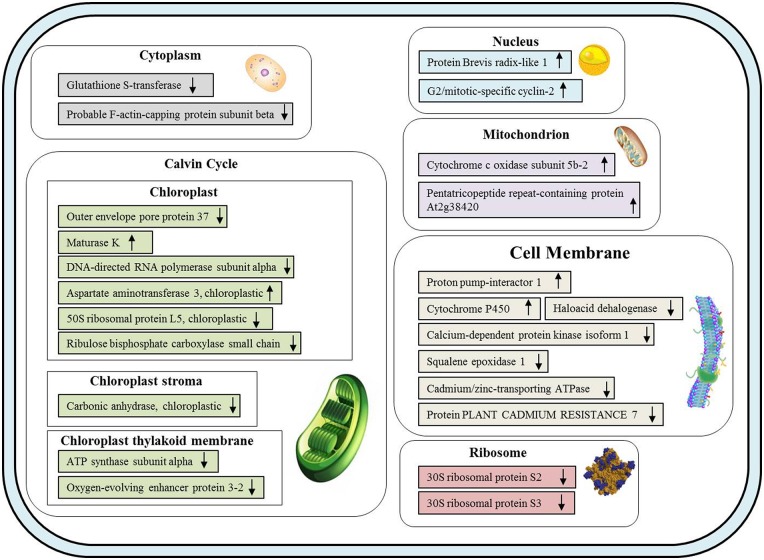
Cadmium (Cd) stress may cause serious morphological and physiological abnormalities in addition to altering the proteome in plants. The present study was performed to explore Cd-induced morpho-physiological alterations and their potential associated mechanisms in Sorghum bicolor leaves at the protein level. Ten-day-old sorghum seedlings were exposed to different concentrations (0, 100, and 150 μM) of CdCl2, and different morpho-physiological responses were recorded. The effects of Cd exposure on protein expression patterns in S. bicolor were investigated using two-dimensional gel electrophoresis (2-DE) in samples derived from the leaves of both control and Cd-treated seedlings. The observed morphological changes revealed that the plants treated with Cd displayed dramatically altered shoot lengths, fresh weights and relative water content. In addition, the concentration of Cd was markedly increased by treatment with Cd, and the amount of Cd taken up by the shoots was significantly and directly correlated with the applied concentration of Cd. Using the 2-DE method, a total of 33 differentially expressed protein spots were analyzed using MALDI-TOF/TOF MS. Of these, treatment with Cd resulted in significant increases in 15 proteins and decreases in 18 proteins. Major changes were absorbed in the levels of proteins known to be involved in carbohydrate metabolism, transcriptional regulation, translation and stress responses. Proteomic results revealed that Cd stress had an inhibitory effect on carbon fixation, ATP production and the regulation of protein synthesis. Our study provides insights into the integrated molecular mechanisms involved in responses to Cd and the effects of Cd on the growth and physiological characteristics of sorghum seedlings. We have aimed to provide a reference describing the mechanisms involved in heavy metal damage to plants.
Conflict of interest statement
Figures
References
-
- Anjum NA, Ahmad I, Mohmood I, Pacheco M, Duarte AC, Pereira E, et al. Modulation of glutathione and its related enzymes in plants’ responses to toxic metals and metalloids-a review. Environmental and Experimental Botany. 2012;75:307–24.
-
- Hossain MA, Hossain MZ, Fujita M. Stress-induced changes of methylglyoxal level and glyoxalase I activity in pumpkin seedlings and cDNA cloning of glyoxalase I gene. Australian Journal of Crop Science. 2009;3(2):53–64.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
