

Host Mitochondrial Association Evolved in the Human Parasite Toxoplasma gondii via Neofunctionalization of a Gene Duplicate
- PMID: 26920761
- PMCID: PMC4858780
- DOI: 10.1534/genetics.115.186270
Host Mitochondrial Association Evolved in the Human Parasite Toxoplasma gondii via Neofunctionalization of a Gene Duplicate
Abstract
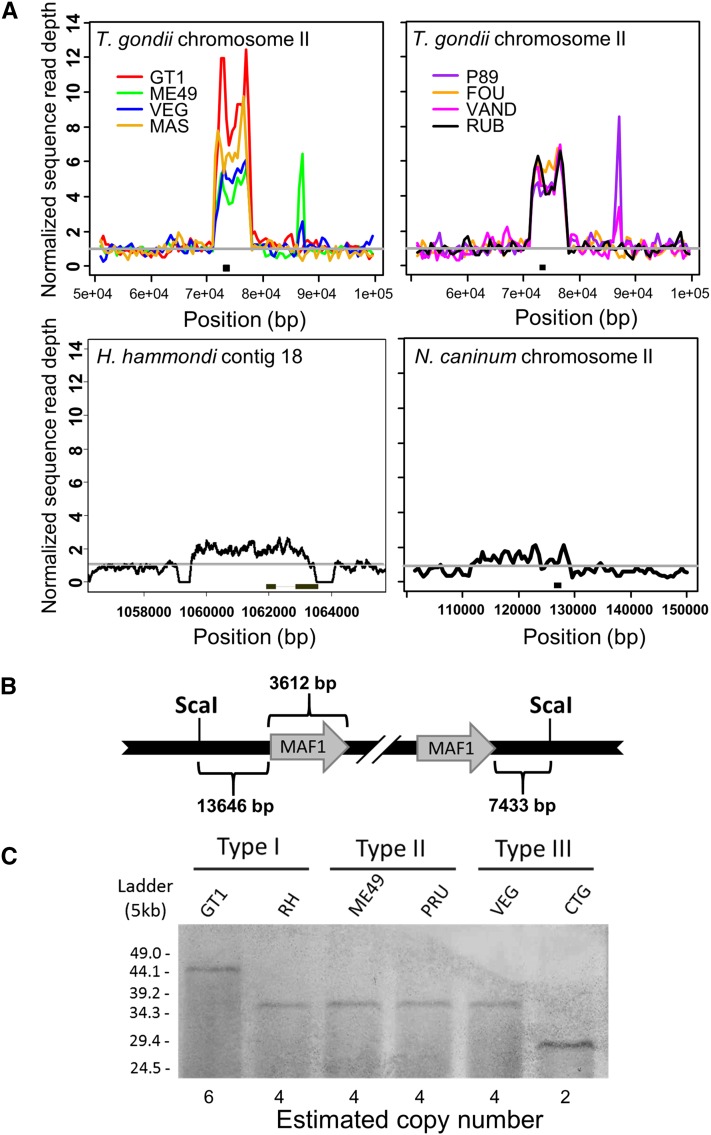
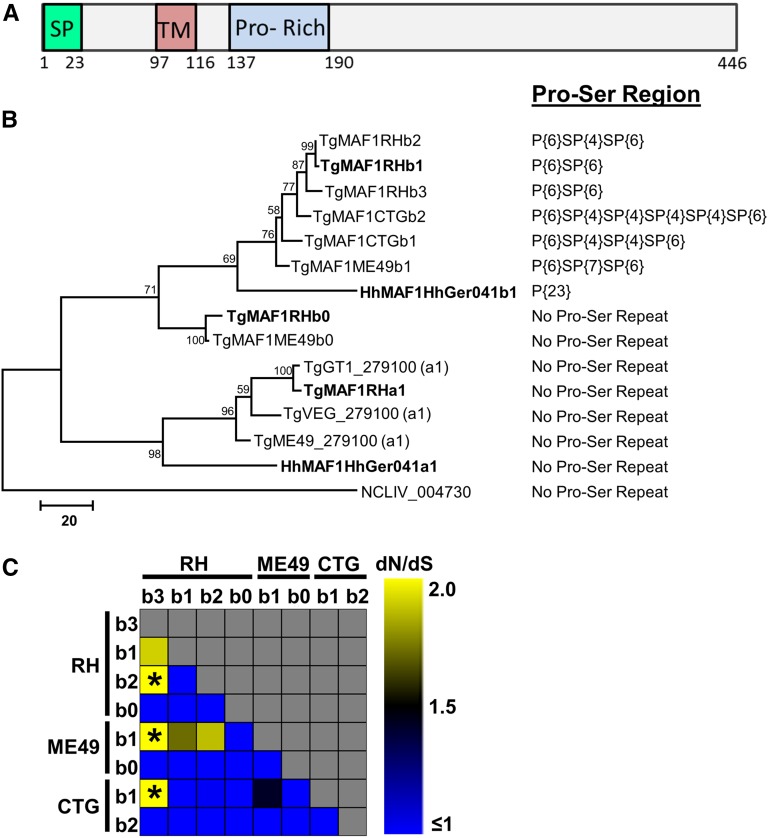
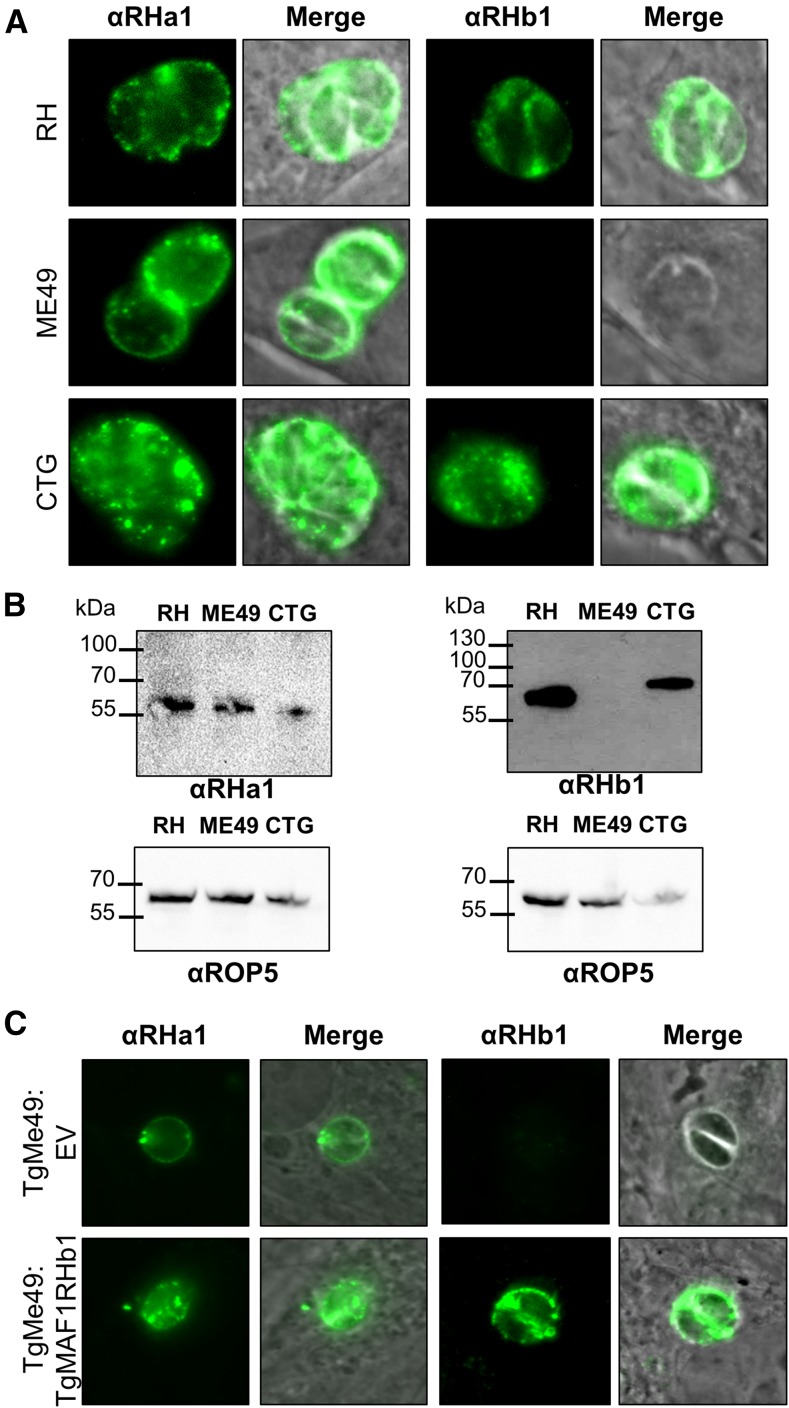
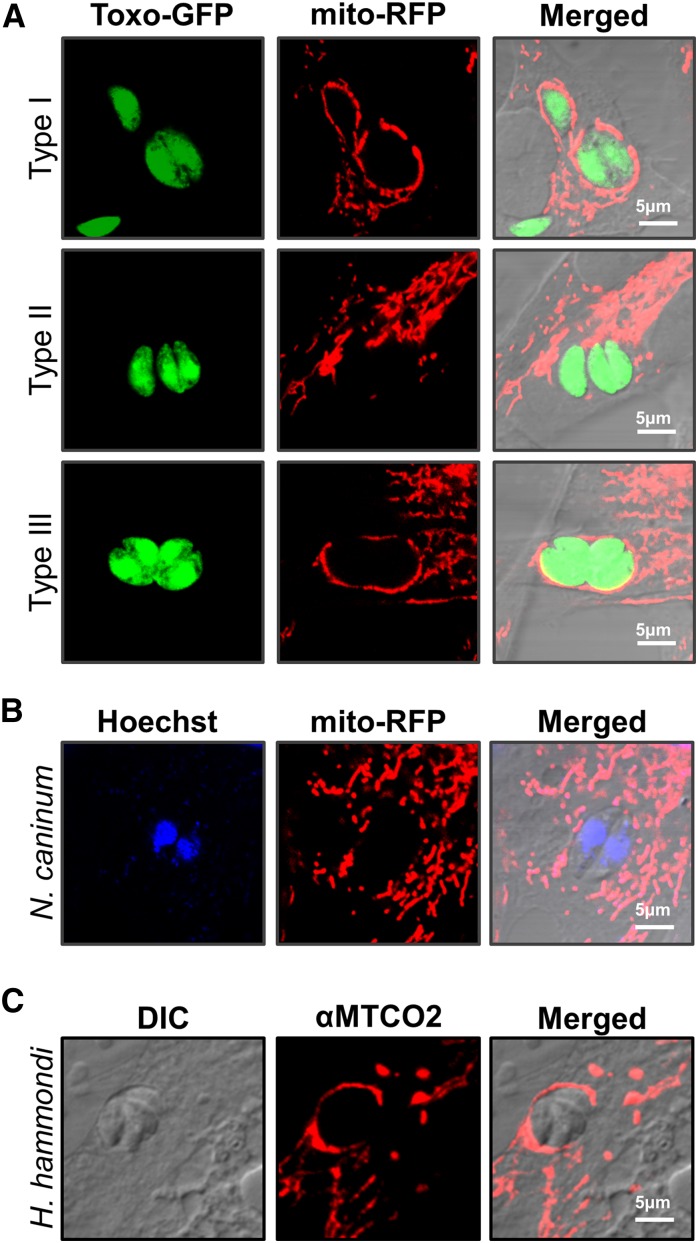
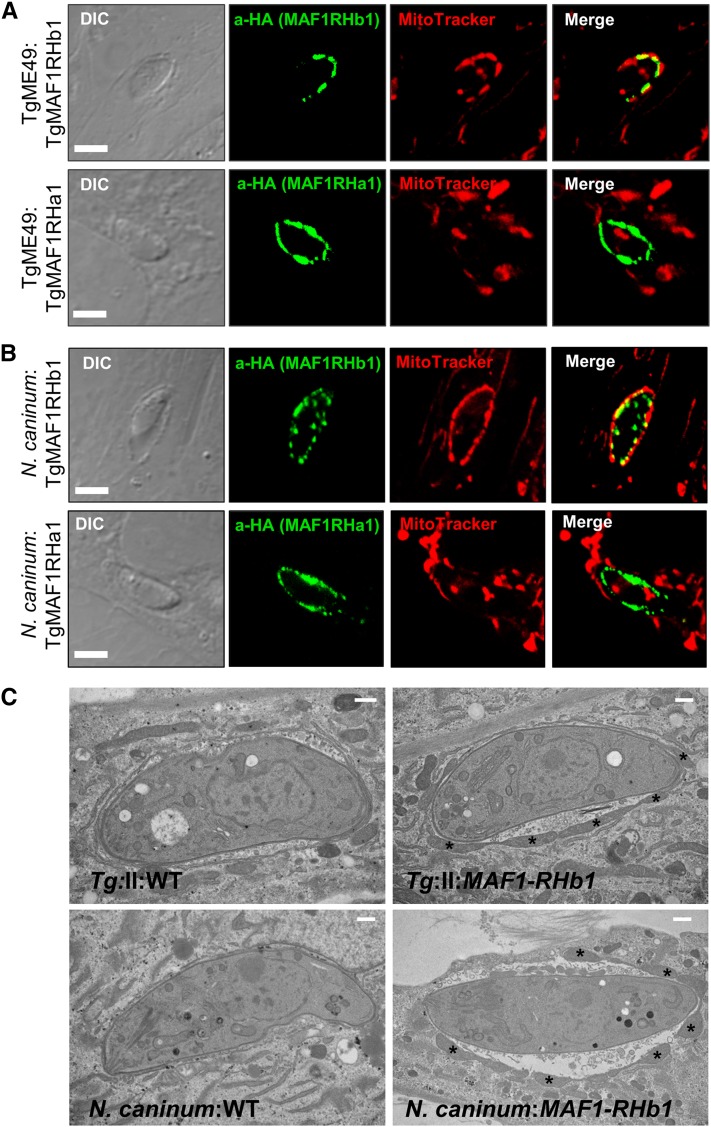
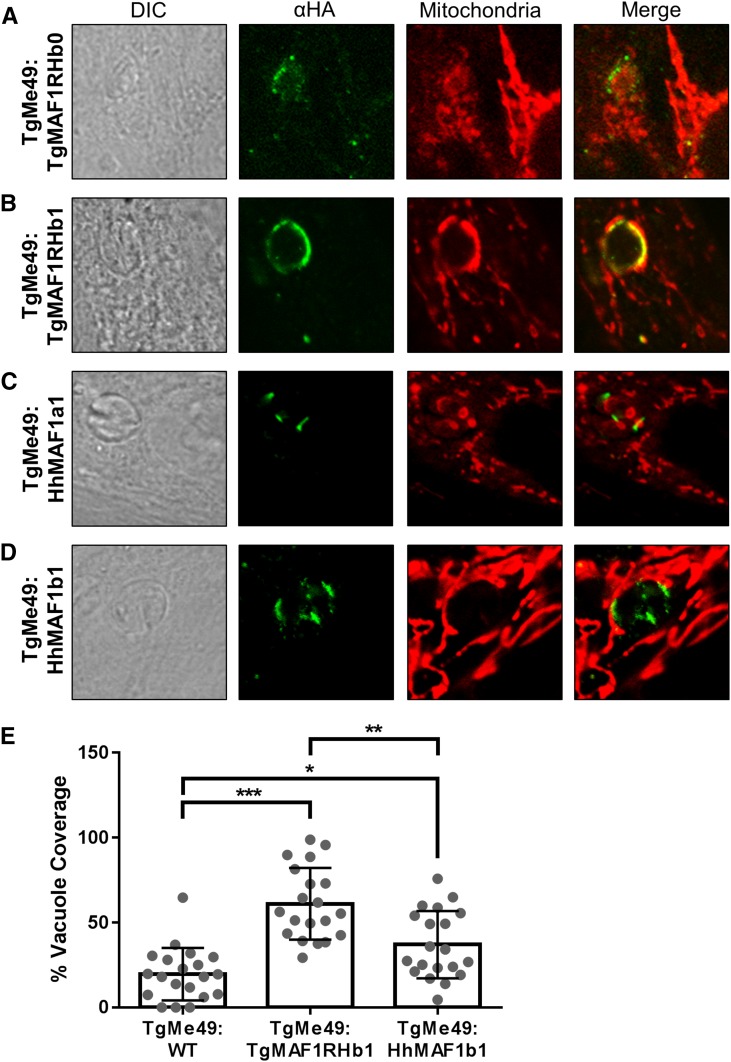
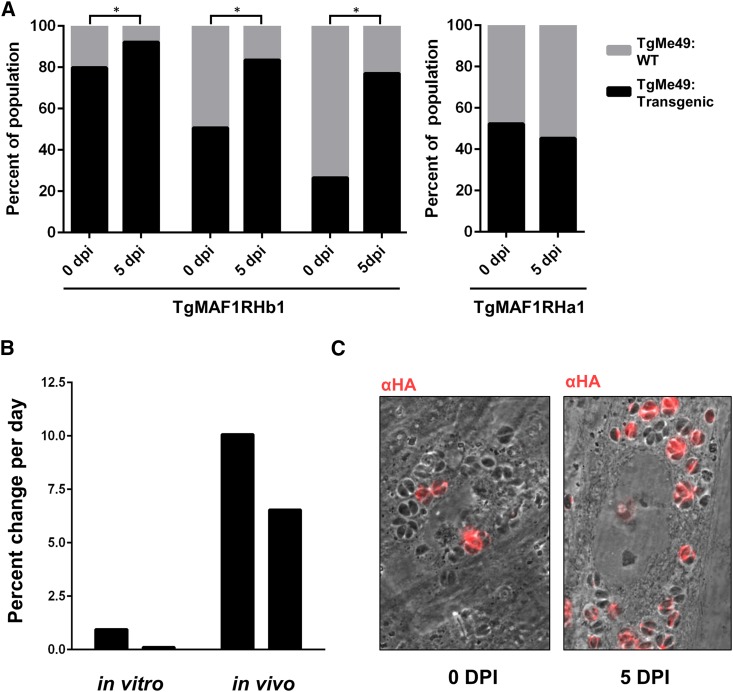
In Toxoplasma gondii, an intracellular parasite of humans and other animals, host mitochondrial association (HMA) is driven by a gene family that encodes multiple mitochondrial association factor 1 (MAF1) proteins. However, the importance of MAF1 gene duplication in the evolution of HMA is not understood, nor is the impact of HMA on parasite biology. Here we used within- and between-species comparative analysis to determine that the MAF1 locus is duplicated in T. gondii and its nearest extant relative Hammondia hammondi, but not another close relative, Neospora caninum Using cross-species complementation, we determined that the MAF1 locus harbors multiple distinct paralogs that differ in their ability to mediate HMA, and that only T. gondii and H. hammondi harbor HMA(+) paralogs. Additionally, we found that exogenous expression of an HMA(+) paralog in T. gondii strains that do not normally exhibit HMA provides a competitive advantage over their wild-type counterparts during a mouse infection. These data indicate that HMA likely evolved by neofunctionalization of a duplicate MAF1 copy in the common ancestor of T. gondii and H. hammondi, and that the neofunctionalized gene duplicate is selectively advantageous.
Keywords: Hammondia hammondi; Neospora caninum; Toxoplasma gondii; gene duplication; neofunctionalization.
Copyright © 2016 by the Genetics Society of America.
Figures
References
-
- Bailey J. A., Eichler E. E., 2006. Primate segmental duplications: crucibles of evolution, diversity and disease. Nat. Rev. Genet. 7: 552–564. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
