

Autoimmune Responses to Soluble Aggregates of Amyloidogenic Proteins Involved in Neurodegenerative Diseases: Overlapping Aggregation Prone and Autoimmunogenic regions
- PMID: 26924748
- PMCID: PMC4770294
- DOI: 10.1038/srep22258
Autoimmune Responses to Soluble Aggregates of Amyloidogenic Proteins Involved in Neurodegenerative Diseases: Overlapping Aggregation Prone and Autoimmunogenic regions
Abstract
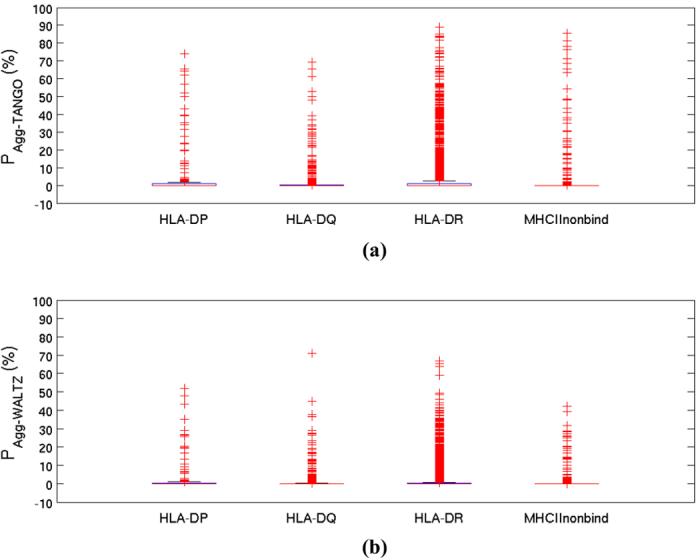
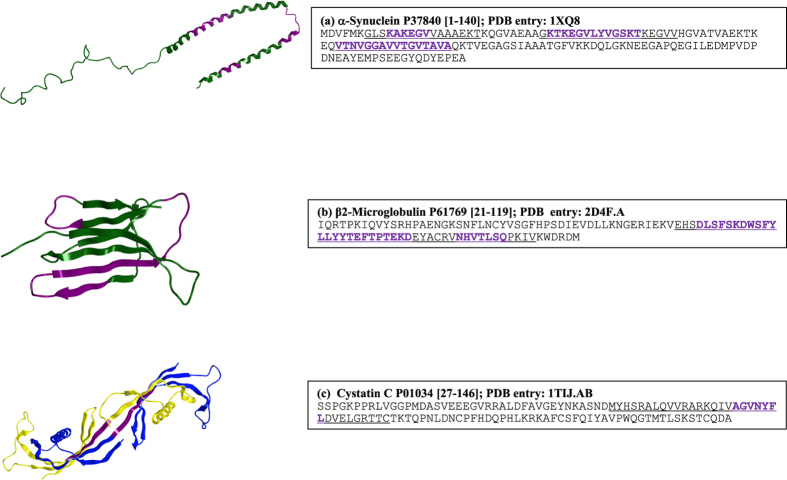
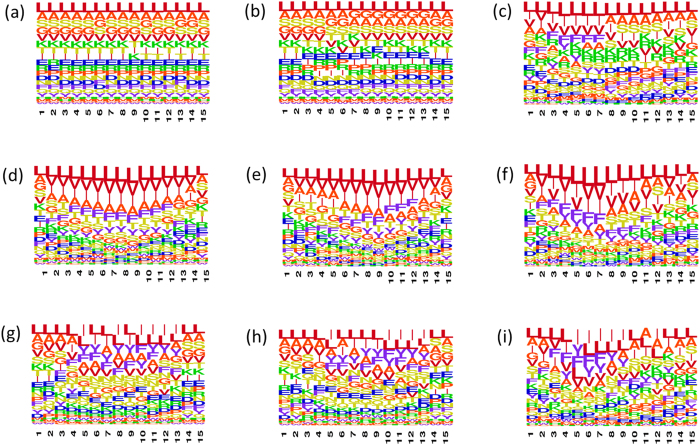
Why do patients suffering from neurodegenerative diseases generate autoantibodies that selectively bind soluble aggregates of amyloidogenic proteins? Presently, molecular basis of interactions between the soluble aggregates and human immune system is unknown. By analyzing sequences of experimentally validated T-cell autoimmune epitopes, aggregating peptides, amyloidogenic proteins and randomly generated peptides, here we report overlapping regions that likely drive aggregation as well as generate autoantibodies against the aggregates. Sequence features, that make short peptides susceptible to aggregation, increase their incidence in human T-cell autoimmune epitopes by 4-6 times. Many epitopes are predicted to be significantly aggregation prone (aggregation propensities ≥10%) and the ones containing experimentally validated aggregating regions are enriched in hydrophobicity by 10-20%. Aggregate morphologies also influence Human Leukocyte Antigen (HLA)--types recognized by the aggregating regions containing epitopes. Most (88%) epitopes that contain amyloid fibril forming regions bind HLA-DR, while majority (63%) of those containing amorphous β-aggregating regions bind HLA-DQ. More than two-thirds (70%) of human amyloidogenic proteins contain overlapping regions that are simultaneously aggregation prone and auto-immunogenic. Such regions help clear soluble aggregates by generating selective autoantibodies against them. This can be harnessed for early diagnosis of proteinopathies and for drug/vaccine design against them.
Figures

Similar articles
-
CPAD, Curated Protein Aggregation Database: A Repository of Manually Curated Experimental Data on Protein and Peptide Aggregation.PLoS One. 2016 Apr 4;11(4):e0152949. doi: 10.1371/journal.pone.0152949. eCollection 2016. PLoS One. 2016. PMID: 27043825 Free PMC article.
-
Relationship between potential aggregation-prone regions and HLA-DR-binding T-cell immune epitopes: implications for rational design of novel and follow-on therapeutic antibodies.J Pharm Sci. 2012 Aug;101(8):2686-701. doi: 10.1002/jps.23169. Epub 2012 May 22. J Pharm Sci. 2012. PMID: 22619033
-
Aggregation prone regions in human proteome: Insights from large-scale data analyses.Proteins. 2017 Jun;85(6):1099-1118. doi: 10.1002/prot.25276. Epub 2017 Mar 24. Proteins. 2017. PMID: 28257595
-
Functional Reciprocity of Amyloids and Antimicrobial Peptides: Rethinking the Role of Supramolecular Assembly in Host Defense, Immune Activation, and Inflammation.Front Immunol. 2020 Jul 31;11:1629. doi: 10.3389/fimmu.2020.01629. eCollection 2020. Front Immunol. 2020. PMID: 32849553 Free PMC article. Review.
-
[Proteinopathies--forms of neurodegenerative disorders with protein aggregation-based pathology].Mol Biol (Mosk). 2012 May-Jun;46(3):402-15. Mol Biol (Mosk). 2012. PMID: 22888630 Review. Russian.
Cited by
-
CPAD, Curated Protein Aggregation Database: A Repository of Manually Curated Experimental Data on Protein and Peptide Aggregation.PLoS One. 2016 Apr 4;11(4):e0152949. doi: 10.1371/journal.pone.0152949. eCollection 2016. PLoS One. 2016. PMID: 27043825 Free PMC article.
-
Targeting Protein Aggregation in ALS.Biomolecules. 2024 Oct 18;14(10):1324. doi: 10.3390/biom14101324. Biomolecules. 2024. PMID: 39456257 Free PMC article. Review.
References
-
- Graus F., Saiz A. & Dalmau J. Antibodies and neuronal autoimmune disorders of the CNS. J Neurol 257, 509–517 (2010). - PubMed
-
- Gruden M. A. et al. Correlation between protective immunity to alpha-synuclein aggregates, oxidative stress and inflammation. Neuroimmunomod 19, 334–342 (2012). - PubMed
-
- Weltzien R. B. & Pachter J. S. Visualization of beta-amyloid peptide (Abeta) phagocytosis by human mononuclear phagocytes: dependency on Abeta aggregate size. J neurosci res 59, 522–527 (2000). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
