

Review
doi: 10.1038/nrg.2015.25.
Epub 2016 Feb 29.
Mechanisms underlying structural variant formation in genomic disorders
Affiliations
- PMID: 26924765
- PMCID: PMC4827625
- DOI: 10.1038/nrg.2015.25
Item in Clipboard
Review
Mechanisms underlying structural variant formation in genomic disorders
Nat Rev Genet.
2016 Apr.
Abstract
With the recent burst of technological developments in genomics, and the clinical implementation of genome-wide assays, our understanding of the molecular basis of genomic disorders, specifically the contribution of structural variation to disease burden, is evolving quickly. Ongoing studies have revealed a ubiquitous role for genome architecture in the formation of structural variants at a given locus, both in DNA recombination-based processes and in replication-based processes. These reports showcase the influence of repeat sequences on genomic stability and structural variant complexity and also highlight the tremendous plasticity and dynamic nature of our genome in evolution, health and disease susceptibility.
Figures
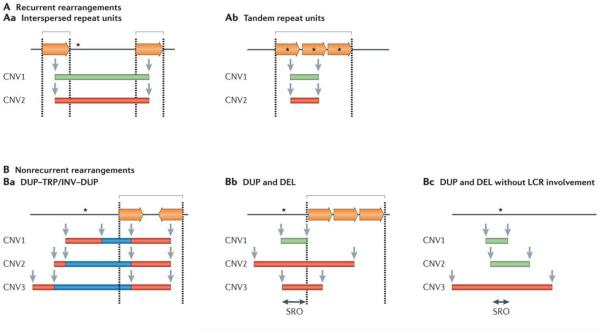
Black lines represent the genomic segments of a given locus. Dosage-sensitive genes that are involved in the rearrangement are represented by black asterisks. Paralogous repeats or low-copy repeats (LCRs) are represented by inverted and directly oriented horizontal orange arrows. Dashed lines indicate LCR regions. Individual genomic copy number variants (CNVs) are represented by colour-matched rectangles (deleted segments (green); duplicated segments (red); and triplicated segments (blue)) and identified as CNV1, CNV2 and CNV3. Locations of CNV breakpoints are indicated by grey vertical arrows. Square brackets denote regions with clustering or grouping of breakpoint junctions. A | Recurrent rearrangements share the same size and genomic content in unrelated individuals. More than 40 non-overlapping genomic disorders are caused by recurrent rearrangements. Aa | In these types of structural variants the duplication or deletion breakpoints cluster within long, highly identical flanking interspersed paralogous repeats (represented by directly oriented horizontal orange arrows) that serve as substrates for nonallelic homologous recombination (NAHR). Ab | Alternatively, recurrent rearrangements can occur within tandem paralogous repeats and affect the copy number of a dosage-sensitive gene that is present within the repeat. For example, ectopic recombination of a tandem repeat consisting of opsin genes at Xq28 can cause red–green colour blindness (OMIM 303900 , OMIM 303800 ) or it can lead to copy number polymorphism at this locus. B | Nonrecurrent rearrangements present a unique size and genomic content at a given locus in unrelated individuals. At least 70 genomic disorders that are caused by nonrecurrent rearrangements have been described. Ba | DUP–TRP/INV–DUP (duplication–inverted triplication–duplication) structures are nonrecurrent but, in some cases, have a limited genomic recurrence with two of four breakpoints mapping to an inverted repeat pair,–. In these cases, the triplicated segments are inverted in relation to the duplications. Bb | Some genomic disorders are characterized by nonrecurrent rearrangements that show breakpoint grouping (rather than clustering as in the recurrent cases discussed above) within paralogous repeats–. Bc | By contrast, some genomic disorders are characterized by nonrecurrent rearrangements without any clustering or grouping of breakpoints–. Green rectangles represent deletions (DEL); red rectangles represent duplications (DUP); blue rectangles represent triplications (TRP). INV, inversion; SRO, smallest region of overlap.
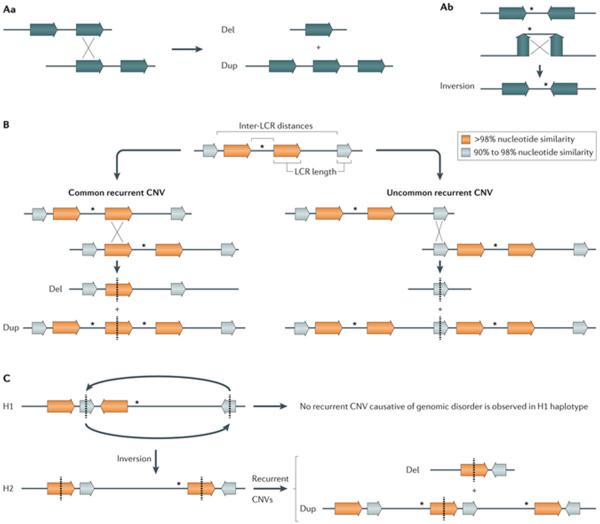
A | Nonallelic homologous recombination (NAHR) between directly oriented repeats generates deletions and duplications (part Aa), whereas NAHR between inverted repeats generates the inversion of the segment in between the repeats (part Ab). B | The rate of NAHR may vary within a particular locus owing to the presence of low-copy repeat (LCR) pairs with distinct susceptibility risk. For example, NAHR rate positively correlates with LCR length and inversely correlates with inter-LCR distance, which can lead to common recurrent and uncommon recurrent copy number variants (CNVs) at a given locus identified in different patients with a genomic disorder (for example, in common recurrent and uncommon recurrent deletions and reciprocal duplications in Smith–Magenis syndrome and Potocki–Lupski syndrome, respectively). C | NAHR rate may vary among individuals with distinct genomic architecture in a given locus or a structural variant haplotype that results in directly oriented LCR. This is exemplified by inverted structural haplotypes that present with distinct risks of undergoing NAHR owing to structural variability in LCR content and orientation. For instance, two divergent structural variant haplotypes, H1 and H2, are observed in 17q21.31 region but deletions that cause Koolen-De Vries syndrome (OMIM 610443 ) occur on chromosomes carrying the H2 haplotype. Dosage-sensitive genes flanked by LCRs are represented by asterisks. LCRs are represented by horizontal colour-matched arrows.
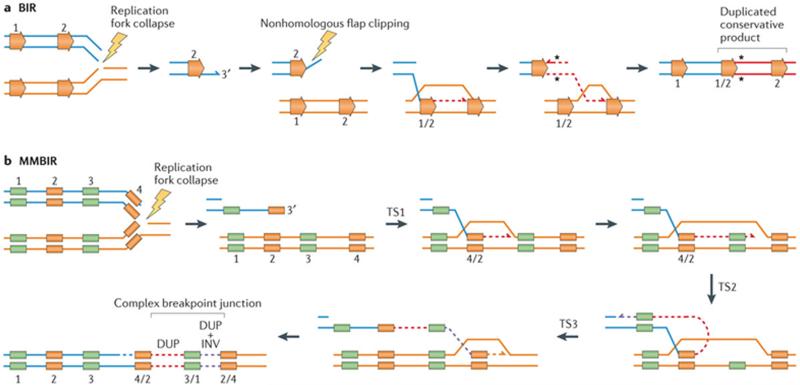
a | Break-induced replication (BIR) can be triggered by a nick on the template strand that may cause stalling or the collapse of the replication fork. Resection of the 5′ end of the broken single-ended, double-stranded DNA (seDNA) molecule exposes a 3′ tail that can invade an allelic (not shown) or a paralogous genomic segment with shared homology (ectopic recombination) to prime replication. Paralogous segments are represented by horizontal orange arrows. The use of ectopic homology to repair broken molecules by BIR can lead to structural variants (for example, duplication (shown here), triplications, deletions and inversions). A conservative mode of repair was recently proposed for BIR that can contribute to perpetuate repair indels or single-nucleotide variant (SNV) (black asterisks) mutations that are acquired during the replicative repair,. b | Microhomology-mediated break-induced replication (MMBIR) can be triggered by a nick on the template strand that may cause stalling or the collapse of the replication fork. Alternatively, MMBIR can also occur by disrupted BIR. Resection of the 5′ end of the broken seDNA molecule exposes a 3′ tail that can anneal to a single-strand DNA sharing microhomology (colour-matched boxes) to prime replication. The initial polymerase extension and replication is carried out by a low processivity polymerase, rendering this repair process prone to undergoing multiple rounds of disengagement and template switches until a fully processive replication fork is established. Short and/or long template switches during repair can generate complexity due to the insertion of templated segments at the rearrangement junction. DUP, duplication; INV, inversion; TS, template switches.
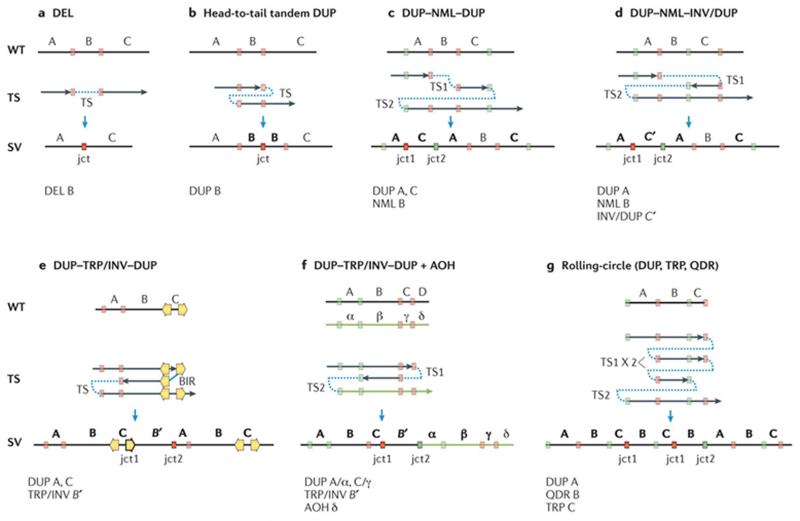
All displayed structural patterns have been identified in one or more genomic disorders (TABLE 1). Top panel: black vertical line indicates a wild-type (WT) genomic segment. Middle panel: black arrows represent segments of DNA generated upon template switches (TSs). Red and green colour-matched boxes represent microhomology regions in WT genomic segment further involved in annealing and resumption of replication during TS. Orange arrows represent inverted low-copy repeats (LCRs) or regions of extended homology that can also be used during TS. Note that formation of any homology/microhomology breakpoint junctions (represented by outlined colour-matched boxes) is accompanied by a relative net reduction of the copy number of those homology/microhomology regions compared to WT regardless of whether the rearrangement results in gain or loss of the flanking genomic material (see microhomology subtractive mode (BOX 1)). Bottom panel: resulting structural variant (SV). Letters (A, B, C, D) represent chromosome alleles. Alleles subject to copy number gains in the resulting SVs are marked as bold letters. Primed letters represent an inverted-oriented genomic segment that originates upon a TS between complementary strands. α, β, γ, δ, represent corresponding homologue alleles. A single TS can generate deletion (DEL) (part a) or a head to tail tandem duplication (DUP) with one junction (jct) (part b). Two TSs can generate distinct complex end products presenting with two breakpoint junctions (jct1 and jct2). DUP–NML–DUP: copy number neutral segment (normal; NML) interspersed between duplications (DUP) (part c). DUP–NML–INV/DUP: inverted interspersed duplications (part d). DUP–TRP/INV–DUP: inverted triplication interspersed with duplications (part e). DUP–TRP/INV–DUP can be generated by break-induced replication (BIR), if inverted repeats are used as substrates for TS, or microhomology-mediated BIR if microhomology is used for TS. DUP–TRP/INV–DUP (part f) can be associated with extended regions of absence of heterozygosity (AOH) if TS occurs between homologous chromosomes (interchromosomal TS). Rolling-circle amplification (part g) can be generated by re-replication (for instance, the generation of two copies of jct1) culminating in the formation of multiple copies of a given segment.
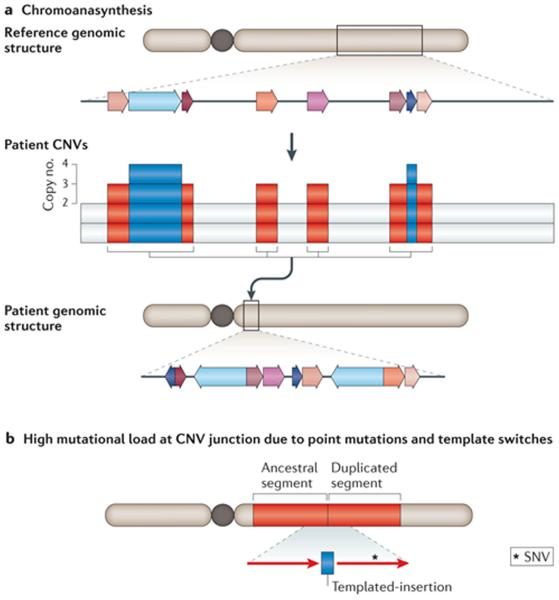
a | Schematic example of copy number variants (CNVs) in multiple, interspersed segments within the same chromosome arm. Chromoanasynthesis in constitutional genomic disorders resembles the phenomenon of chromosome catastrophes (that is, chromothripsis) initially described based on the analyses of hundreds of cancer genome sequences and in different types of cancers. The chromosomal region involved in chromoanasynthesis is denoted by a grey box; local genomic segments involved are represented by coloured arrows. Array comparative genomic hybridization (aCGH) reveals genomic segments with copy number variation. The rearranged chromosome is shown on which the extra copy number segments (represented by colour-matched arrows) are inserted together in a new chromosomal position with respect to the reference genome. This clustering of CNVs in a new position is a hallmark of chromoanasynthesis. b | Increased mutational load can be observed at the breakpoint junctions of CNVs generated by replication-based mechanisms (RBMs). SNVs include point mutations and short insertions/short deletions (indels). Red rectangles represent the duplicated segment and red arrows represent the orientation of the duplicated segments.
References
-
- Lupski JR. Genomic disorders: structural features of the genome can lead to DNA rearrangements and human disease traits. Trends Genet. 1998;14:417–422. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
